
International Journal of
Reproductive Biomedicine
Tue, Jul 14, 2026
[Archive]
Volume 6, Issue 2 (7-2008)
IJRM 2008, 6(2): 39-0 |
Back to browse issues page

Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Khosrowbeygi A, Zargham N, Farzadi L. Fatty acid composition in normozoospermic, asthenozoopermic, asthenoteratozoospermic and oligoasthenoteratozoospermic ejaculates. IJRM 2008; 6 (2) :39
URL: http://ijrm.ir/article-1-98-en.html
URL: http://ijrm.ir/article-1-98-en.html


1- School of Medicine, Lorestan University of Medical Sciences, Khoramabad, Lorestan, Iran
2- Drug Applied Research Center, Tabriz University of Medical Sciences, Tabriz, Iran ,zarghami@tbzmed.ac.ir
3- Department of Obstetrics and Gynecology, Tabriz University of Medical Sciences, Tabriz, Iran
2- Drug Applied Research Center, Tabriz University of Medical Sciences, Tabriz, Iran ,
3- Department of Obstetrics and Gynecology, Tabriz University of Medical Sciences, Tabriz, Iran
Keywords: Sperm, Fatty acid, Asthenozoospermia, Asthenoteratozoospermia, Oligoasthenoteratozoospermia.
Full-Text [PDF 89 kb]
(691 Downloads)
| Abstract (HTML) (3560 Views)
Full-Text: (444 Views)
Introduction
The membrane structure of spermatozoa plays a pivotal role for successful fertilization, since both the acrosome reaction and sperm-oocyte fusion are membrane-associated events (1-3). The lipids of the spermatozoa membrane are important for the fluidity and flexibility of spermatozoa.
However, spermatozoa’s lipids are the main substrates for peroxidation, which may provoke severe functional disorder of sperm (3-7).
The findings on the sperm fatty acid composition are controversial and the pathological groups that were studied are limited. Zalata et al (1) observed that sperm fatty acid composition of asthenozoospermic and oligozoospermic samples differ from that of normozoospermic individuals. Their study showed that levels of stearic acid are higher in both case groups. On the other hand, levels of docosahexaenoic acid (DHA) are lower in both asthenozoospermic and oligozoospermic samples than in normozoospermic males. Zalata et al study also showed that DHA levels correlate positively with sperm motility and morphology. While Conquer et al (8) study showed that levels of oleic acid are significantly higher in the sperm of asthenozoospermic samples compared with normozoospermic men. On the other hand, levels of DHA are significantly lower in the sperm of asthenozoospermic individuals. Conquer et al also obseved that DHA levels of spermatozoa is correlated positively with sperm motility and concentration. In addition Gulaya et al (9) study showed that fatty acid composition of total sperm of infertile men is altered compared with fertile males. They observed that in infertile males the levels of DHA are dramatically lowered. Gulaya et al study also showed that there is a significant positive correlation between DHA levels and sperm motility. Calamera et al (10), observed that sperm from normozoospermic individuals have lower unsaturated fatty acid content than those from asthenozoospermic individuals. In another study Aksoy et al (11) showed that polyunsaturated fatty acids (PUFA) are lower in spermatozoa from asthenozoospermic, oligo-asthenozoospermic and oligozoospermic men than those from normozoospermic males. Similarly Tavilani et al (12) observed that spermatozoa from asthenozoospermic samples have higher levels of saturated fatty acids and lower levels of PUFA compared with normozoospermic men.
The aim of this study was to investigate the fatty acid composition of spermatozoa in men with asthenozoospermia, asthenoteratozoospermia and oligoasthenoteratozoospermia compared to normozoospermic males.
Materials and methods
Sampling
A cross-sectional study with a simple random sampling was designed. Following Institutional Review board approval the semen samples were collected from subjects. The semen samples were collected from males undergoing infertility screening from September 2005 to April 2006. All specimens were collected into sterile plastic containers by masturbation after an abstinence period of 3-5 days, at the clinical andrology laboratory at IVF Center of Tabriz University of Medical Sciences and were analyzed within 1h of collection. All specimens were allowed at least 30 min for liquefaction to occur, semen analysis was performed to measure sperm concentration, normal sperm morphology and percentage of sperm motility using Sperm Quality Analyzer IIC (SQA IIC, United Medical Systems Inc, Santa Ana, CA, USA) (13, 14). Samples with a leukocyte concentration >106/ml of ejaculate were excluded from this study. The differentiation of leukocytes and spermatogonic cells was performed by the myeloperoxidase staining (LeucoScreen; FertiPro) (1). The criteria for sperm normality were as follows: sperm concentration ≥20×106/ml of ejaculate, progressively motile sperms ≥50% and normal sperm morphology ≥30% (13, 14). The patients consisted of 51 men with seminal parameters abnormalities. The patients were grouped into asthenozoospermic (n=15), asthenoteratozoospermic (n=21) and oligoastheno-teratozoospermic (n=15). The patients were compared with 21 males with normozoospermia. Samples with sperm concentration ≥20×106/ml of ejaculate, motility <50% and morphology ≥30% were considered asthenozoospermic. Samples with sperm concentration ≥20×106/ml of ejaculate, motility <50%, and morphology <30% were considered asthenoteratozoospermic. Samples with sperm concentration <20×106/ml of ejaculate, motility <50% and morphology <30% were considered oligoasthenoteratozoospermic. Liquefied semen samples were centrifuged at 15000 g for 10 minutes (8). The pellet sperms were then frozen at -80ºC until further analysis.
Sperm fatty acid analysis
Sperm fatty acid analysis was performed using capillary gas chromatography (10). Sperms were diluted with one volume of Dulbecco’s phosphate buffered saline (PBS) and centrifuged at 800 g for 8 min. This centrifugation step was repeated, and the pellet resuspended in 500 μl of PBS. Lipids were extracted by liquid-liquid extraction with six volumes of chloroform: methanol (2:1, v/v), centrifuged at 800g for 3 min, and the resulting lower phase was aspirated. An aliquot of 30 μl of a 0.25 mg/ml solution of heptadecanoic acid (17:0) in chloroform: methanol (2:1, v/v) was added as an internal standard to this solution and then dried. Fatty acids from extracts were transmethylated by boron trifluoride-methanol (15%) reagent (Merck, Darmstadt, Germany). Dry extracts were resuspended in 500 μl of boron trifluoride-methanol reagent, vortexed and incubated at 60ºC for 30 min. The 500 μl of water and 1 ml of hexane were added to this solution, vortexed and the hexane upper phase was transferred to a new glass tube and dried. The residue was redissolved in 100 μl hexane, of which 1 μl was injected in a GC-2010 Gas Chromatograph (Shimadzu, Japan). An OMEGAWAX 250 capillary column (Supelco, USA) of 30-m length and 0.25-mm internal diameter was used. Initial temperature was 170ºC and final temperature 275ºC at 7ºC/min. Flame ionization detector (FID) temperature was 300ºC. Fatty acid methyl ester peaks were identified by comparison of retention times of standards and quantified in comparison with the internal standard detector response. Fatty acids concentration was expressed as n-mole percent.
Statistical analysis
Based on a pilot study, using an α value of 0.05 and a β value of 0.2 (80% power), the minimum sample size required was 15 samples per group. Differences between groups were assessed using Mann-Whitney U test and Kruskal-Wallis test. Coefficients of correlation were calculated using Spearman’s correlation analysis. All hypothesis tests were two-tailed with statistical significance assessed at the p-value<0.05 level with 95% confidence intervals. The data were expressed as the mean ± SEM.
Results
Seminal parameters of the subjects are reported in Table I. Sperm fatty acid profile (palmitic acid, stearic acid, oleic acid, linoleic acid, arachidonic acid, and DHA) of patients and normozoospermic subjects was shown in Table II. The most abundant saturated fatty acids and PUFAs in the normozoospermic, asthenozoospermic, astheno-teratozoospermic, and oligoasthenoterato-zoospermic subjects were palmitic acid and DHA, respectively. Levels of DHA were slightly decreased in asthenozoospermic and asthenoterato-zoospermic samples compared with normo-zoospermic individuals. In asthenoteratozoo-spermic group oleic acid showed a significant increase compared with normozoospermic samples (p=0.007). Levels of stearic acid and oleic acid were significantly higher in oligoasthenoterato-zoospermic subjects than those from normozoospermic males (p=0.009, p=0.04, respectively). On the other hand, levels of DHA and arachidonic acid were significantly lower in the sperms of oligoasthenoteratozoospermic males (p=0.03). Linolenic acid was not detected in any samples.
The correlation between sperm fatty acid levels and sperm motility and morphology was assessed in total samples (n=72). Sperm motility showed an inverse correlation with levels of stearic acid (r=-0.38, p=0.001) and oleic acid (r=-0.33, p=0.001). On the other hand, sperm motility showed a positive correlation with arachidonic acid (r=0.25, p=0.04) and DHA (r=0.29, p=0.01) levels. Sperm morphology showed a significant negative correlation with levels of stearic acid (r =-0.39, p=0.001) and oleic acid (r=-0.34, p=0.004) while a positive correlation was observed with levels of arachidonic acid (r=0.25, p=0.04) and DHA (r=0.31, p=0.009).

Discussion
The most relevant findings of this study were (I) a significant increase in levels of stearic acid and oleic acid and decrease in levels of DHA in sperms of patients compared to normozoospermic males and (II) a significant positive correlation between sperm motility and normal morphology and levels of sperm total DHA.
The high levels of palmitic acid and stearic acid as saturated fatty acid, oleic acid as monounsaturated fatty acid, and DHA as PUFA in spermatozoa, as suggested in the literature (1, 15-19), are supported by our results. In contrary to the literature (1, 8-12), linolenic acid was not detected in our study. Various factors such as the methods of sperm preparation and fatty acid analysis may contribute to this difference.
Zalata et al (1) observed that sperm fatty acid composition of asthenozoospermic and oligozoospermic individuals differ from normozoospermic individuals. In their study, sperm were separated on a Percoll gradient (in order to separate the most active sperm from the least active sperm in a given individual) and recovered from the 90% Percoll and 47% Percoll fractions. Zalata et al study showed that levels of DHA are decreased in both asthenozoospermic and oligozoospermic samples compared to normozoospermic males. Our results were nearly similar to Zalata et al finding (Table II).
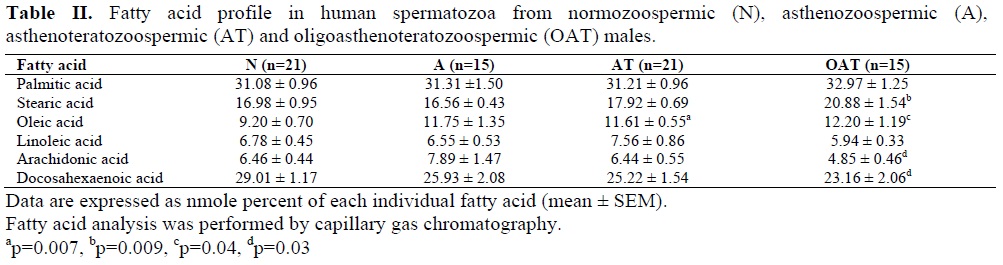
Conquer et al (8) study showed that levels of oleic acid are significantly higher in the sperm of asthenozoospermic vs. normozoospermic men while levels of DHA are significantly lower. In our study levels of oleic acid were slightly increased and levels of DHA were slightly decreased (Table II). Similar to Conquer et al Study, in our study levels of palmitic acid, stearic acid, linoleic acid, and arachidonic acid did not alter statistically significant in asthenozoospermic specimens compared with normozoospermic men (Table II).
Gulaya et al (9) observed that in infertile males the levels of DHA are significantly lowered. Our result was similar to Gulaya et al study. Aksoy et al (11) showed that sperm fatty acid composition of asthenozoospermic, oligoasthenozoospermic and oligozoospermic men differ from that of normozoospermic males. Their study showed that DHA is significantly lower in asthenozoospermic, oligoasthenozoospermic and oligozoospermic samples compared to normozoospermic males. Our results about DHA were nearly similar to Aksoy et al finding (Table II). Tavilani et al (12) observed that spermatozoa from asthenozoospermic samples have higher levels of saturated fatty acid and lower levels of PUFAs compared to normozoospermic men. They found that levels of palmitic acid and stearic acid are significantly increased while linoleic acid and DHA are dramatically increased. In contrary to Tavilani et al study, in our study levels of palmitic acid, stearic acid and linoleic acid in asthenozoospermic samples did not show any significant difference from normozoospermic men and levels of DHA were slightly decreased. In contrary to our results and the literature (1, 8, 9,11, 12), Calamera et al (10) found that that sperm from normozoospermic individuals have lower unsaturated fatty acid content than sperm from asthenozoospermic individuals . They also assessed superoxide dismutase (SOD) content of spermatozoa which is increased in normozoospermic individuals. Therefore, Calamera et al concluded that spermatozoa from normozoospermic samples are less susceptible to Reactive oxygen species (ROS) -induced peroxidative damage.
Defective sperm function is the most common cause of infertility. Oxidative stress has long been believed to influence male reproductive function and the main targets for ROS and DNA and membrane lipids. Immature spermatozoa with abnormal morphology and cytoplasmic retention are the major source of ROS production in semen as has been reported by Gil-Guzman et al study (3). They showed that there is a direct and significant correlation between the average ROS levels and the rate of abnormal forms in semen. Mammalian spermatozoa membranes are rich in PUFA. It has been shown that the oxidation potential of PUFA is several times greater than that of monounsaturated fatty acids (11, 12). Therefore, lower content of PUFAs in spermatozoa of oligoasthenoteratozoospermic subjects may be result of breakdown of them due to high levels of ROS in these samples (12).
We found significant correlation between sperm DHA concentrations and sperm motility and normal morphology. Our correlation results are in agreement with results of Zalata et al (1), Conquer et al (8), Gulaya et al (9), Aksoy et al (11) and Tavilani et al (12). These findings may indicate that PUFAs are related to normal sperm structure, production and function (11).
In a conclusion, impaired sperm function can originate from the disorder of sperm lipid metabolism. Low levels of PUFAs in spermatozoa of oligoasthenoteratozoospermic subjects may be result of breakdown of them. Further studies with higher sample size are required to prove this conclusion.
The membrane structure of spermatozoa plays a pivotal role for successful fertilization, since both the acrosome reaction and sperm-oocyte fusion are membrane-associated events (1-3). The lipids of the spermatozoa membrane are important for the fluidity and flexibility of spermatozoa.
However, spermatozoa’s lipids are the main substrates for peroxidation, which may provoke severe functional disorder of sperm (3-7).
The findings on the sperm fatty acid composition are controversial and the pathological groups that were studied are limited. Zalata et al (1) observed that sperm fatty acid composition of asthenozoospermic and oligozoospermic samples differ from that of normozoospermic individuals. Their study showed that levels of stearic acid are higher in both case groups. On the other hand, levels of docosahexaenoic acid (DHA) are lower in both asthenozoospermic and oligozoospermic samples than in normozoospermic males. Zalata et al study also showed that DHA levels correlate positively with sperm motility and morphology. While Conquer et al (8) study showed that levels of oleic acid are significantly higher in the sperm of asthenozoospermic samples compared with normozoospermic men. On the other hand, levels of DHA are significantly lower in the sperm of asthenozoospermic individuals. Conquer et al also obseved that DHA levels of spermatozoa is correlated positively with sperm motility and concentration. In addition Gulaya et al (9) study showed that fatty acid composition of total sperm of infertile men is altered compared with fertile males. They observed that in infertile males the levels of DHA are dramatically lowered. Gulaya et al study also showed that there is a significant positive correlation between DHA levels and sperm motility. Calamera et al (10), observed that sperm from normozoospermic individuals have lower unsaturated fatty acid content than those from asthenozoospermic individuals. In another study Aksoy et al (11) showed that polyunsaturated fatty acids (PUFA) are lower in spermatozoa from asthenozoospermic, oligo-asthenozoospermic and oligozoospermic men than those from normozoospermic males. Similarly Tavilani et al (12) observed that spermatozoa from asthenozoospermic samples have higher levels of saturated fatty acids and lower levels of PUFA compared with normozoospermic men.
The aim of this study was to investigate the fatty acid composition of spermatozoa in men with asthenozoospermia, asthenoteratozoospermia and oligoasthenoteratozoospermia compared to normozoospermic males.
Materials and methods
Sampling
A cross-sectional study with a simple random sampling was designed. Following Institutional Review board approval the semen samples were collected from subjects. The semen samples were collected from males undergoing infertility screening from September 2005 to April 2006. All specimens were collected into sterile plastic containers by masturbation after an abstinence period of 3-5 days, at the clinical andrology laboratory at IVF Center of Tabriz University of Medical Sciences and were analyzed within 1h of collection. All specimens were allowed at least 30 min for liquefaction to occur, semen analysis was performed to measure sperm concentration, normal sperm morphology and percentage of sperm motility using Sperm Quality Analyzer IIC (SQA IIC, United Medical Systems Inc, Santa Ana, CA, USA) (13, 14). Samples with a leukocyte concentration >106/ml of ejaculate were excluded from this study. The differentiation of leukocytes and spermatogonic cells was performed by the myeloperoxidase staining (LeucoScreen; FertiPro) (1). The criteria for sperm normality were as follows: sperm concentration ≥20×106/ml of ejaculate, progressively motile sperms ≥50% and normal sperm morphology ≥30% (13, 14). The patients consisted of 51 men with seminal parameters abnormalities. The patients were grouped into asthenozoospermic (n=15), asthenoteratozoospermic (n=21) and oligoastheno-teratozoospermic (n=15). The patients were compared with 21 males with normozoospermia. Samples with sperm concentration ≥20×106/ml of ejaculate, motility <50% and morphology ≥30% were considered asthenozoospermic. Samples with sperm concentration ≥20×106/ml of ejaculate, motility <50%, and morphology <30% were considered asthenoteratozoospermic. Samples with sperm concentration <20×106/ml of ejaculate, motility <50% and morphology <30% were considered oligoasthenoteratozoospermic. Liquefied semen samples were centrifuged at 15000 g for 10 minutes (8). The pellet sperms were then frozen at -80ºC until further analysis.
Sperm fatty acid analysis
Sperm fatty acid analysis was performed using capillary gas chromatography (10). Sperms were diluted with one volume of Dulbecco’s phosphate buffered saline (PBS) and centrifuged at 800 g for 8 min. This centrifugation step was repeated, and the pellet resuspended in 500 μl of PBS. Lipids were extracted by liquid-liquid extraction with six volumes of chloroform: methanol (2:1, v/v), centrifuged at 800g for 3 min, and the resulting lower phase was aspirated. An aliquot of 30 μl of a 0.25 mg/ml solution of heptadecanoic acid (17:0) in chloroform: methanol (2:1, v/v) was added as an internal standard to this solution and then dried. Fatty acids from extracts were transmethylated by boron trifluoride-methanol (15%) reagent (Merck, Darmstadt, Germany). Dry extracts were resuspended in 500 μl of boron trifluoride-methanol reagent, vortexed and incubated at 60ºC for 30 min. The 500 μl of water and 1 ml of hexane were added to this solution, vortexed and the hexane upper phase was transferred to a new glass tube and dried. The residue was redissolved in 100 μl hexane, of which 1 μl was injected in a GC-2010 Gas Chromatograph (Shimadzu, Japan). An OMEGAWAX 250 capillary column (Supelco, USA) of 30-m length and 0.25-mm internal diameter was used. Initial temperature was 170ºC and final temperature 275ºC at 7ºC/min. Flame ionization detector (FID) temperature was 300ºC. Fatty acid methyl ester peaks were identified by comparison of retention times of standards and quantified in comparison with the internal standard detector response. Fatty acids concentration was expressed as n-mole percent.
Statistical analysis
Based on a pilot study, using an α value of 0.05 and a β value of 0.2 (80% power), the minimum sample size required was 15 samples per group. Differences between groups were assessed using Mann-Whitney U test and Kruskal-Wallis test. Coefficients of correlation were calculated using Spearman’s correlation analysis. All hypothesis tests were two-tailed with statistical significance assessed at the p-value<0.05 level with 95% confidence intervals. The data were expressed as the mean ± SEM.
Results
Seminal parameters of the subjects are reported in Table I. Sperm fatty acid profile (palmitic acid, stearic acid, oleic acid, linoleic acid, arachidonic acid, and DHA) of patients and normozoospermic subjects was shown in Table II. The most abundant saturated fatty acids and PUFAs in the normozoospermic, asthenozoospermic, astheno-teratozoospermic, and oligoasthenoterato-zoospermic subjects were palmitic acid and DHA, respectively. Levels of DHA were slightly decreased in asthenozoospermic and asthenoterato-zoospermic samples compared with normo-zoospermic individuals. In asthenoteratozoo-spermic group oleic acid showed a significant increase compared with normozoospermic samples (p=0.007). Levels of stearic acid and oleic acid were significantly higher in oligoasthenoterato-zoospermic subjects than those from normozoospermic males (p=0.009, p=0.04, respectively). On the other hand, levels of DHA and arachidonic acid were significantly lower in the sperms of oligoasthenoteratozoospermic males (p=0.03). Linolenic acid was not detected in any samples.
The correlation between sperm fatty acid levels and sperm motility and morphology was assessed in total samples (n=72). Sperm motility showed an inverse correlation with levels of stearic acid (r=-0.38, p=0.001) and oleic acid (r=-0.33, p=0.001). On the other hand, sperm motility showed a positive correlation with arachidonic acid (r=0.25, p=0.04) and DHA (r=0.29, p=0.01) levels. Sperm morphology showed a significant negative correlation with levels of stearic acid (r =-0.39, p=0.001) and oleic acid (r=-0.34, p=0.004) while a positive correlation was observed with levels of arachidonic acid (r=0.25, p=0.04) and DHA (r=0.31, p=0.009).
Discussion
The most relevant findings of this study were (I) a significant increase in levels of stearic acid and oleic acid and decrease in levels of DHA in sperms of patients compared to normozoospermic males and (II) a significant positive correlation between sperm motility and normal morphology and levels of sperm total DHA.
The high levels of palmitic acid and stearic acid as saturated fatty acid, oleic acid as monounsaturated fatty acid, and DHA as PUFA in spermatozoa, as suggested in the literature (1, 15-19), are supported by our results. In contrary to the literature (1, 8-12), linolenic acid was not detected in our study. Various factors such as the methods of sperm preparation and fatty acid analysis may contribute to this difference.
Zalata et al (1) observed that sperm fatty acid composition of asthenozoospermic and oligozoospermic individuals differ from normozoospermic individuals. In their study, sperm were separated on a Percoll gradient (in order to separate the most active sperm from the least active sperm in a given individual) and recovered from the 90% Percoll and 47% Percoll fractions. Zalata et al study showed that levels of DHA are decreased in both asthenozoospermic and oligozoospermic samples compared to normozoospermic males. Our results were nearly similar to Zalata et al finding (Table II).
Conquer et al (8) study showed that levels of oleic acid are significantly higher in the sperm of asthenozoospermic vs. normozoospermic men while levels of DHA are significantly lower. In our study levels of oleic acid were slightly increased and levels of DHA were slightly decreased (Table II). Similar to Conquer et al Study, in our study levels of palmitic acid, stearic acid, linoleic acid, and arachidonic acid did not alter statistically significant in asthenozoospermic specimens compared with normozoospermic men (Table II).
Gulaya et al (9) observed that in infertile males the levels of DHA are significantly lowered. Our result was similar to Gulaya et al study. Aksoy et al (11) showed that sperm fatty acid composition of asthenozoospermic, oligoasthenozoospermic and oligozoospermic men differ from that of normozoospermic males. Their study showed that DHA is significantly lower in asthenozoospermic, oligoasthenozoospermic and oligozoospermic samples compared to normozoospermic males. Our results about DHA were nearly similar to Aksoy et al finding (Table II). Tavilani et al (12) observed that spermatozoa from asthenozoospermic samples have higher levels of saturated fatty acid and lower levels of PUFAs compared to normozoospermic men. They found that levels of palmitic acid and stearic acid are significantly increased while linoleic acid and DHA are dramatically increased. In contrary to Tavilani et al study, in our study levels of palmitic acid, stearic acid and linoleic acid in asthenozoospermic samples did not show any significant difference from normozoospermic men and levels of DHA were slightly decreased. In contrary to our results and the literature (1, 8, 9,11, 12), Calamera et al (10) found that that sperm from normozoospermic individuals have lower unsaturated fatty acid content than sperm from asthenozoospermic individuals . They also assessed superoxide dismutase (SOD) content of spermatozoa which is increased in normozoospermic individuals. Therefore, Calamera et al concluded that spermatozoa from normozoospermic samples are less susceptible to Reactive oxygen species (ROS) -induced peroxidative damage.
Defective sperm function is the most common cause of infertility. Oxidative stress has long been believed to influence male reproductive function and the main targets for ROS and DNA and membrane lipids. Immature spermatozoa with abnormal morphology and cytoplasmic retention are the major source of ROS production in semen as has been reported by Gil-Guzman et al study (3). They showed that there is a direct and significant correlation between the average ROS levels and the rate of abnormal forms in semen. Mammalian spermatozoa membranes are rich in PUFA. It has been shown that the oxidation potential of PUFA is several times greater than that of monounsaturated fatty acids (11, 12). Therefore, lower content of PUFAs in spermatozoa of oligoasthenoteratozoospermic subjects may be result of breakdown of them due to high levels of ROS in these samples (12).
We found significant correlation between sperm DHA concentrations and sperm motility and normal morphology. Our correlation results are in agreement with results of Zalata et al (1), Conquer et al (8), Gulaya et al (9), Aksoy et al (11) and Tavilani et al (12). These findings may indicate that PUFAs are related to normal sperm structure, production and function (11).
In a conclusion, impaired sperm function can originate from the disorder of sperm lipid metabolism. Low levels of PUFAs in spermatozoa of oligoasthenoteratozoospermic subjects may be result of breakdown of them. Further studies with higher sample size are required to prove this conclusion.
Acknowledgement
This research was supported by a grant from the Drug Applied Research Center of Tabriz University of Medical Sciences. We thank Dr. Yaghoob Assadi (University of Science and Technology, Tehran, Iran) for his skillful technical assistance. The authors are grateful to Tabriz IVF center for their cooperation in this research.
Type of Study: Original Article |
References
1. Zalata AA, Christophe AB, Depuydt CE, Schoonjans F, Comhaire FH. The fatty acid composition of phospholipids of spermatozoa from infertile patients. Mol Hum Reprod 1998; 4: 111-118. [DOI:10.1093/molehr/4.2.111]
2. Ollero M, Powers RD, Alvarez JG. Variation of docosahexaenoic acid content in subsets of human spermatozoa at different stages of maturation: implications for sperm lipoperoxidative damage. Mol Reprod Dev 2000; 55: 326-334.
https://doi.org/10.1002/(SICI)1098-2795(200003)55:3<326::AID-MRD11>3.0.CO;2-A [DOI:10.1002/(SICI)1098-2795(200003)55:33.0.CO;2-A]
3. Gil-Guzman E, Ollero M, Lopez MC, Sharma RK, Alvarez JG, Thomas Jr AJ , et al. Differential production of reactive oxygen species by subsets of human spermatozoa at different stages of maturation. Hum Reprod 2001; 16: 1922-1930. [DOI:10.1093/humrep/16.9.1922]
4. Zarghami N, Khosrowbeygi A. Evaluation of lipid peroxidation as an indirect measure of oxidative stress in seminal plasma. Iranian J Reprod Med 2004; 2: 34-39.
5. Khosrowbeygi A, Zarghami N, Deldar Y. Correlation between sperm quality parameters and seminal plasma antioxidants status. Iranian J Reprod Med 2004; 2: 58-64.
6. Agarwal A, Prabakaran S. Oxidative stress and antioxidants in male infertility: a difficult balance. Iranian J Reprod Med 2005; 3: 1-8.
7. Khosrowbeygi A, Zarghami N. Levels of oxidative stress biomarkers in seminal plasma and their relationship with seminal parameters. BMC Clin Pathol 2007; 7: 6. [DOI:10.1186/1472-6890-7-6]
8. Conquer JA, Martin JB, Tummon I, Watson L, Tekpetey F. Fatty acid analysis of blood serum, seminal plasma, and spermatozoa of normozoospermic vs. asthenozoospermic males. Lipids 1999; 34: 793-799. [DOI:10.1007/s11745-999-0425-1]
9. Gulaya NM, Margitich VM, Govseeva NM, Klimashevsky VM, Gorpynchenko II, Boyko MI. Phospholipid composition of human sperm and seminal plasma in relation to sperm fertility. Arch Androl 2001; 46: 169-175. [DOI:10.1080/01485010151096405]
10. Calamera J, Buffone M, Ollero M, Alvarez J, Doncel GF. Superoxide dismutase content and fatty acid composition in subsets of human spermatozoa from normozoospermic, asthenozoospermic, and polyzoospermic semen samples. Mol Reprod Dev 2003; 66: 422-430. [DOI:10.1002/mrd.10368]
11. Aksoy Y, Aksoy H, Altinkaynak K, Aydin HR, Ozkan A. Sperm fatty acid composition in subfertile men. Prostaglandins, Leukotrienes and Essential Fatty Acids 2006; 75: 75-79. [DOI:10.1016/j.plefa.2006.06.002]
12. Tavilani H, Doosti M, Abdi K, Vaisiraygani A, Joshaghani H. Decreased polyunsaturated and increased saturated fatty acid concentration in spermatozoa from asthenozoospermic males as compared with normozoospermic males. Andrologia 2006; 38: 173-178. [DOI:10.1111/j.1439-0272.2006.00735.x]
13. Shibahara H, Suzuki T, Obara H, Hirano Y, Onagawa T, Taneichi A, et al. Accuracy of the normal sperm morphology value by Sperm Quality Analyzer IIC: comparison with the strict criteria. Int J Androl 2002; 25: 45-48. [DOI:10.1046/j.1365-2605.2002.00322.x]
14. Suzuki T, Shibahara H, Tsunoda H, Hirano Y, Taneichi A, Obara H, et al. Comparison of the Sperm Quality Analyzer IIC variables with the computer-aided sperm analysis estimates. Int J Androl 2002; 25: 49-54. [DOI:10.1046/j.1365-2605.2002.00324.x]
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
