
International Journal of
Reproductive Biomedicine
Sun, May 31, 2026
[Archive]
Volume 6, Issue 4 (7-2008)
IJRM 2008, 6(4): 143-0 |
Back to browse issues page

Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Absalan F, Movahedin M, Mowla S J. Assessment of testis histological changes and sperm parameters in experimentally–induced unilateral and bilateral cryptorchid mouse model. IJRM 2008; 6 (4) :143-0
URL: http://ijrm.ir/article-1-114-en.html
URL: http://ijrm.ir/article-1-114-en.html


1- Department of Anatomical Sciences, School of Medical Sciences, Tehran, Iran
2- Department of Anatomical Sciences, School of Medical Sciences, Tehran, Iran ,mansoure@modares.ac.ir
3- Department of Genetics, School of Basic Sciences, Tarbiat Modares University, Tehran, Iran
2- Department of Anatomical Sciences, School of Medical Sciences, Tehran, Iran ,
3- Department of Genetics, School of Basic Sciences, Tarbiat Modares University, Tehran, Iran
Full-Text [PDF 121 kb]
(812 Downloads)
| Abstract (HTML) (3656 Views)
Full-Text: (522 Views)
Introduction
It was shown that mammalian testis must descend from the abdominal cavity for normal development to occur, and that the elevated testicular temperature disrupts spermatogenesis and causes infertility (1).
Testicular maldescend is a common urological disorder. In full-term infants, the incidence is 3.4% to 5.8% and at the time of puberty, 0.8%- 1.8% of boys remain cryptorchid (2).
The abdominal positioning of cryptorchid testis is associated with disruption of germinal epithelium (3). In addition to degeneration of the seminiferous epithelium, accumulation of lipid in sertoli cells, local dilations of the interacellular spaces between sertoli cells junctions and modifications in the interstitial tissue have been reported (4).
Damber et al showed that blood flow to the cryptorchid testis is reduced despite the fact that the cryptorchid testis contains a greater percentage and number of blood vessels than the normal testis (5).
Leydig cell hypertrophy and hyperplasia are accompanied by an increase in the cytoplasmic organelles involved in steroidogenesis, such as smooth endoplasmic reticulum and mitochondria (6). By contrast, Hall et al reported that steriodogenic functions of the somatic cells (Leydig and Sertoli cells) appear to be normal when exposed to the core body temperature (7). The elevated testicular temperature affects the normal functioning of all major cell types (Germ, Leydig and Sertoli cells) to some extend (8). Also heat-induced germ cell degeneration is usually accompanied by alteration in Sertoli cell morphology and function, a decrease in testes fluid, and increase in serum FSH with no changes in serum Testosterone and LH levels (9). More recently, a number of studies have documented that high temperatures induced an increased synthesis of several proteins and a decreased synthesis of many others.
The effect of temperature on testicular function, especially sperm motility and sperm viability are unclear however, several enzymatic activities are affected and the lipid composition of plasma membranes is modified (10). Besides, testicular stem cell enrichment has been widely used to investigate the biology of these cells in addition to the production of transgenic animals, germ cell transplantation and restoration of fertility in various species. The potential model for enrichment is cryptorchidism (11). In this study, we are reporting the effects of prolong cryptorchidism on sperm parameters and testis microstructure of unilateral and bilateral experimental cryptorchidism. We believe that the present model is a suitable tool for enrichment of spermatogonial stem cells also it is useful for the treatment of cryptorchidism and further biological research in to spermatogenes.
Materials and methods
Animals
Immature NMRI mice at under 2 months of age (Weigh 7-12 gr) were purchased from Razi Vaccine & Serum Research Institute (Karaj, Iran). Mice were anesthetized with an injection of 1.6ml/kg of a mixture of Ketamine and xylezine. A cut was made along the skin in the upper abdominal region and the adipose tissue of the caput epididymis was sutured to the inner peritoneal wall, pushing the testes in to the abdomen.
The first experimental group mice were unilaterally cryptorchidized and in the second experimental group, mice were bilaterally cryptorchidized. Experimental and control animals were sacrificed by cervical dislocation, 2, 4, 6 and 8 weeks after surgery. Following cervical dislocation left testis of unilateral groups and both testes of bilateral groups and epididymis were removed, the testes were weighed and fixed in Bouin,s fixative, dehydrated and embedded in paraffin.
Then, 5-µm serial microscopic sections were prepared and at least 5 slides from each testis stained with hematoxylin and eosin for histological assessment.
In each experiment, at least 5 animals were prepared and analyzed (13). All animal experimentation protocols were approved by the Institutional Animal Care and Use Committee of Tarbiat Modarres University (Tehran, Iran).
Sperm parameters assessment
The epididymis was placed in 1 ml PBS (Ph 7.4) and minced into small pieces before being incubated at 37°C for 30min. Sperm parameters were monitored by light microscopy. Sperms viability was assessed by determining the percentage of sperms excluding vital dye (25% eosin solution). Briefly, 7 µl of eosin solution was added to 20 µl of cell suspension after incubation and mixed thoroughly. Motility of sperm was also assessed by determining the percentage of motile sperm (14). Finally sperm numbers was also calculated with a hemocytometer count and compared in unilateral, bilateral, sham and control groups.
Morphometrical analysis of testis
For each testis, in 100 randomly selected tubular profiles that were round or nearly round, the diameters of tubules and epithelium thickness were measured under light microscopy. Volume density of spermatogonial cells, spermatocytes and spermatids in seminiferous tubules were determined.
The location and morphology of the cells within the seminiferous tubules were used to identity them. An estimate of each parameter was performed by examining 20 fields in 5 histological sections from each testis (13).
Statistical analysis
The results were analyzed by performing ANOVA and Tukey,s tests, with p<0.05 considered as statistically significant. The mean and standard deviation (SD) was also calculated for each value.
Results
Analyses of sperm parameters in epididymis following creating unilateral and bilateral cryptorchidism mouse model
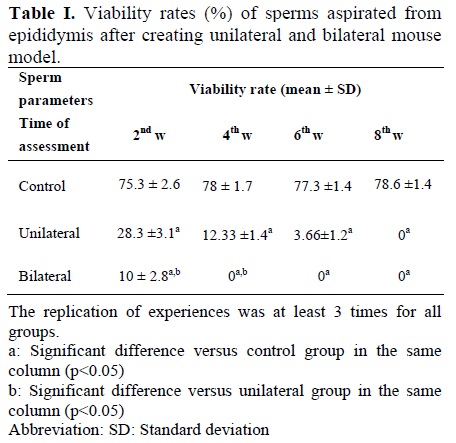
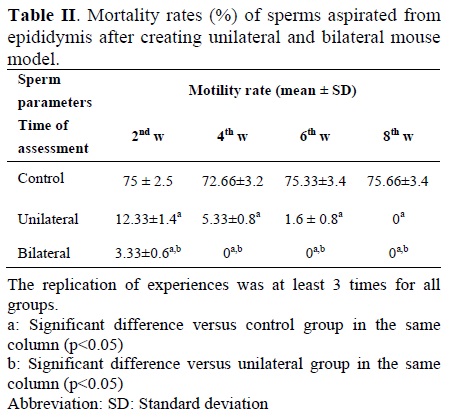
There was no significant difference between the result of sham and control groups. As the results show, there was a significant decline in sperm viability and motility rates (Table I and II) as well as a decline in sperms counts (Table III) following surgery. The viability of sperms decreased in all weeks after surgery, however, only in 2nd and 4th week after surgery there was significant difference between unilateral and bilateral groups. Sperm viability was not observed after 4th week in bilateral group but this situation occurs in unilateral group after 8th week (Table I). Similar observation was seen on sperm motility after surgery. However, the effect of heat on sperm motility was more severe. The significant difference between unilateral and bilateral groups only observed 2 weeks after surgery (Table II). The number of sperms per ml aspirated from one epididym was determined by hemocytometer counts. As presented in Table III, in all times after surgery the number of sperms decreased significantly in epididymis of experimental groups compared with control group but there was no significant difference between unilateral and bilateral groups (Table III). In addition, we showed decline in all sperm parameters after heat exposure especially in bilateral group.
Analyses of morphological changes of mouse testis following creating unilateral and bilateral cryptorchid mouse model
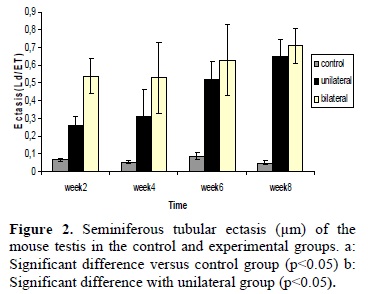
As the data show, the values of the testicular weight in unilateral and bilateral groups were significantly less than those in the control group. These decrease observed from 2 week in both experimental groups but significant difference between these groups only observed in 4th week after surgery (figure 1). Histolological examination of the testis in control and experimentally groups revealed that tubular ectasis (rate of seminiferous tubules diameter comparing to the seminiferous epithelium thickness) was increased in experimental groups in time dependent manner. This difference started from 2 week after surgery between control and experimental groups (figure 2). The testes were histologically examined by H&E staining (figure 3). Compared to the testes in the control group, in cryptorchid model, testicular tissues were degenerated. Spermatogonia, spermatocyte and round spermatids were observed,

but the differentiated cells after them, elongated spermatids or spermatozoa were not detected in the experimental groups. Numbers of spermatogonia increased after 8th week post surgery in bilateral group, while absence or sloughing of germ cells were not found in all experimental animals (Table IV). Based on these results the bilateral cryptorchid model is a useful tool for enrichment of spermatogonial stem cells.



Discussion
Most mammals have a scrotum, and it is well established that the mild hyperthermia leads to infertility (15,16). As a sequence, the effects of local heating on the animal testis have been widely studied (17). In the current study we are reporting the effects of prolong heating caused by surgery on sperm parameters and testis microstructure of mouse testis also we compared this effects in unilateral and bilateral experimental cryptorchidism mouse model.
One of the findings in this study was the decrease in tsetis wieght and in the number of spermatozoa in testis and epididymis in experimental mouse toward the end of experiment, associated with a significant deterioration in the histology. This finding indicates that degeneration of spermatocytes and spermatids occures after cryptorchidism is in agreement with Chowdhury and Steinberger (18,19) also by Collis and Lacy (20). By 2 weeks post operation, the mean weight of the testes in the bilateral cryptorchid mouse had diminished about %50 of the weight in the control group. Many studies suggested that germ cell loss after heat exposure caused this decrease in testicular weight (21). Longquan (2006) demonsterated that experimentally induced cryptorchidism in adult male rats caused a decrease in the percentage of motile spermatozoa within 3 days post operation (22). The present study demonstrated that experimentally induced cryptorchidism in immature mouse caused the epidydimal percentage of sperm number, motility and viability rates decreased significantly from 2 week postoperation in both experimental groups. Motility and viability rates decline in bilateral experimental group compared with unilateral group.
The seminiferous tubules in the cryptorchid testes were similar in shape in cross section to those of the control testes, but their diameter showed progressive reduction with the increasing duration of cryptprchidism. The volume density measurements showed that the seminiferous tubule lumen is increased after cryptorchidism. Degenaration of germ cells resulted in the increase in the tubule lumen (4,6). We showed tubular ectasis increased after surgery and this subject was more prominent in bilateral groups compared with unilateral groups probably due to rapid degeneration of differentiated germ cells in bilateral groups. Many investigators showed significant decreased diameters of seminiferous tubules and thickness of germinal epithelium in cryptorchid testis (23).
During spermatogenesis, the spermatogonial stem cells (SSC) which undergo self-renewal throughout adult life of the animal and produces progeny cells, undergo the complicated differentiation steps. As in other rare stem cell populations, enrichment strategies are required to study SSC biology. It has been demonstrated that during testis development, the total number of SSC per testis increases 39-fold from newborn to adult (24). The number of SSC also appears to increase during the regeneration of spermatogenesis following cytotoxic damage (25, 26). Either testis hyperthermia or cryptorchidism may be used to increase the percentage of SSC invivo. Although these experimental conditions cause depletion of more differentiated germ cells, they do not appear to affect SSC survival and biological activity, thus resulting in an increase in the ratio of SSC vs. the total number of germ cells (27). An effect of heat on spermatogonia in mice had already been reported by Reid, who performed studies on the repoplution of seminiferous tubules after irridiation (28). Differentiation of spermatogonia has also been reported to be arrested in cryptorchid mice (29) and heating the testes of sheep caused a reduction in the production of A1 spermatogonia 20 days later, with no effect on the numbers of A0 spermatogonia (30). Miralgia and Hayashi (31) reported a decrease in the percentage of tubules with spermatogonia from 100 to 89% at day 15 after heating the testes of rats to 43ºC for 30 min. We showed spermatocytes and spermatids are major cells which were affected also spermatogonial number increased after 2 week postoperation and this increase is noticeable in bilateral group compared with unilateral group. In addition, experimentally induced bilateral cryptorchidism provides a useful model for enrichment of spermatogonial stem cells.
According to previous study, experimentally induced unilateral cryptorchidism provides a useful model for study of the role of local factors regulating testis cell apoptosis this suggested that apoptotic cell death induced by bilateral cryptorchidism might be affected by changes in systemic factors. The increase of apoptosis in male germ cell after unilateral cryptorchidism is presumably regulated by local testicular factors (32). Also in this research, the differences indicated between unilateral and bilateral groups maybe arise from this subject.
We believe that the present model is a suitable tool for enrichment of spermatogonial stem cells; also it is useful for treatment of cryptorchidism and further biological research in spermatogenesis.
Acknowledgments
This work was supported by a research grant from Tarbiat Modares University.
We highly thank Shahram Pour Beiranvand and Saeedeh Ebrahimi (Anatomical Sciences Department, Faculty of Medical Sciences, Tarbiat Modares University) for their technical assistant.
It was shown that mammalian testis must descend from the abdominal cavity for normal development to occur, and that the elevated testicular temperature disrupts spermatogenesis and causes infertility (1).
Testicular maldescend is a common urological disorder. In full-term infants, the incidence is 3.4% to 5.8% and at the time of puberty, 0.8%- 1.8% of boys remain cryptorchid (2).
The abdominal positioning of cryptorchid testis is associated with disruption of germinal epithelium (3). In addition to degeneration of the seminiferous epithelium, accumulation of lipid in sertoli cells, local dilations of the interacellular spaces between sertoli cells junctions and modifications in the interstitial tissue have been reported (4).
Damber et al showed that blood flow to the cryptorchid testis is reduced despite the fact that the cryptorchid testis contains a greater percentage and number of blood vessels than the normal testis (5).
Leydig cell hypertrophy and hyperplasia are accompanied by an increase in the cytoplasmic organelles involved in steroidogenesis, such as smooth endoplasmic reticulum and mitochondria (6). By contrast, Hall et al reported that steriodogenic functions of the somatic cells (Leydig and Sertoli cells) appear to be normal when exposed to the core body temperature (7). The elevated testicular temperature affects the normal functioning of all major cell types (Germ, Leydig and Sertoli cells) to some extend (8). Also heat-induced germ cell degeneration is usually accompanied by alteration in Sertoli cell morphology and function, a decrease in testes fluid, and increase in serum FSH with no changes in serum Testosterone and LH levels (9). More recently, a number of studies have documented that high temperatures induced an increased synthesis of several proteins and a decreased synthesis of many others.
The effect of temperature on testicular function, especially sperm motility and sperm viability are unclear however, several enzymatic activities are affected and the lipid composition of plasma membranes is modified (10). Besides, testicular stem cell enrichment has been widely used to investigate the biology of these cells in addition to the production of transgenic animals, germ cell transplantation and restoration of fertility in various species. The potential model for enrichment is cryptorchidism (11). In this study, we are reporting the effects of prolong cryptorchidism on sperm parameters and testis microstructure of unilateral and bilateral experimental cryptorchidism. We believe that the present model is a suitable tool for enrichment of spermatogonial stem cells also it is useful for the treatment of cryptorchidism and further biological research in to spermatogenes.
Materials and methods
Animals
Immature NMRI mice at under 2 months of age (Weigh 7-12 gr) were purchased from Razi Vaccine & Serum Research Institute (Karaj, Iran). Mice were anesthetized with an injection of 1.6ml/kg of a mixture of Ketamine and xylezine. A cut was made along the skin in the upper abdominal region and the adipose tissue of the caput epididymis was sutured to the inner peritoneal wall, pushing the testes in to the abdomen.
The first experimental group mice were unilaterally cryptorchidized and in the second experimental group, mice were bilaterally cryptorchidized. Experimental and control animals were sacrificed by cervical dislocation, 2, 4, 6 and 8 weeks after surgery. Following cervical dislocation left testis of unilateral groups and both testes of bilateral groups and epididymis were removed, the testes were weighed and fixed in Bouin,s fixative, dehydrated and embedded in paraffin.
Then, 5-µm serial microscopic sections were prepared and at least 5 slides from each testis stained with hematoxylin and eosin for histological assessment.
In each experiment, at least 5 animals were prepared and analyzed (13). All animal experimentation protocols were approved by the Institutional Animal Care and Use Committee of Tarbiat Modarres University (Tehran, Iran).
Sperm parameters assessment
The epididymis was placed in 1 ml PBS (Ph 7.4) and minced into small pieces before being incubated at 37°C for 30min. Sperm parameters were monitored by light microscopy. Sperms viability was assessed by determining the percentage of sperms excluding vital dye (25% eosin solution). Briefly, 7 µl of eosin solution was added to 20 µl of cell suspension after incubation and mixed thoroughly. Motility of sperm was also assessed by determining the percentage of motile sperm (14). Finally sperm numbers was also calculated with a hemocytometer count and compared in unilateral, bilateral, sham and control groups.
Morphometrical analysis of testis
For each testis, in 100 randomly selected tubular profiles that were round or nearly round, the diameters of tubules and epithelium thickness were measured under light microscopy. Volume density of spermatogonial cells, spermatocytes and spermatids in seminiferous tubules were determined.
The location and morphology of the cells within the seminiferous tubules were used to identity them. An estimate of each parameter was performed by examining 20 fields in 5 histological sections from each testis (13).
Statistical analysis
The results were analyzed by performing ANOVA and Tukey,s tests, with p<0.05 considered as statistically significant. The mean and standard deviation (SD) was also calculated for each value.
Results
Analyses of sperm parameters in epididymis following creating unilateral and bilateral cryptorchidism mouse model
There was no significant difference between the result of sham and control groups. As the results show, there was a significant decline in sperm viability and motility rates (Table I and II) as well as a decline in sperms counts (Table III) following surgery. The viability of sperms decreased in all weeks after surgery, however, only in 2nd and 4th week after surgery there was significant difference between unilateral and bilateral groups. Sperm viability was not observed after 4th week in bilateral group but this situation occurs in unilateral group after 8th week (Table I). Similar observation was seen on sperm motility after surgery. However, the effect of heat on sperm motility was more severe. The significant difference between unilateral and bilateral groups only observed 2 weeks after surgery (Table II). The number of sperms per ml aspirated from one epididym was determined by hemocytometer counts. As presented in Table III, in all times after surgery the number of sperms decreased significantly in epididymis of experimental groups compared with control group but there was no significant difference between unilateral and bilateral groups (Table III). In addition, we showed decline in all sperm parameters after heat exposure especially in bilateral group.
Analyses of morphological changes of mouse testis following creating unilateral and bilateral cryptorchid mouse model
As the data show, the values of the testicular weight in unilateral and bilateral groups were significantly less than those in the control group. These decrease observed from 2 week in both experimental groups but significant difference between these groups only observed in 4th week after surgery (figure 1). Histolological examination of the testis in control and experimentally groups revealed that tubular ectasis (rate of seminiferous tubules diameter comparing to the seminiferous epithelium thickness) was increased in experimental groups in time dependent manner. This difference started from 2 week after surgery between control and experimental groups (figure 2). The testes were histologically examined by H&E staining (figure 3). Compared to the testes in the control group, in cryptorchid model, testicular tissues were degenerated. Spermatogonia, spermatocyte and round spermatids were observed,
but the differentiated cells after them, elongated spermatids or spermatozoa were not detected in the experimental groups. Numbers of spermatogonia increased after 8th week post surgery in bilateral group, while absence or sloughing of germ cells were not found in all experimental animals (Table IV). Based on these results the bilateral cryptorchid model is a useful tool for enrichment of spermatogonial stem cells.
Discussion
Most mammals have a scrotum, and it is well established that the mild hyperthermia leads to infertility (15,16). As a sequence, the effects of local heating on the animal testis have been widely studied (17). In the current study we are reporting the effects of prolong heating caused by surgery on sperm parameters and testis microstructure of mouse testis also we compared this effects in unilateral and bilateral experimental cryptorchidism mouse model.
One of the findings in this study was the decrease in tsetis wieght and in the number of spermatozoa in testis and epididymis in experimental mouse toward the end of experiment, associated with a significant deterioration in the histology. This finding indicates that degeneration of spermatocytes and spermatids occures after cryptorchidism is in agreement with Chowdhury and Steinberger (18,19) also by Collis and Lacy (20). By 2 weeks post operation, the mean weight of the testes in the bilateral cryptorchid mouse had diminished about %50 of the weight in the control group. Many studies suggested that germ cell loss after heat exposure caused this decrease in testicular weight (21). Longquan (2006) demonsterated that experimentally induced cryptorchidism in adult male rats caused a decrease in the percentage of motile spermatozoa within 3 days post operation (22). The present study demonstrated that experimentally induced cryptorchidism in immature mouse caused the epidydimal percentage of sperm number, motility and viability rates decreased significantly from 2 week postoperation in both experimental groups. Motility and viability rates decline in bilateral experimental group compared with unilateral group.
The seminiferous tubules in the cryptorchid testes were similar in shape in cross section to those of the control testes, but their diameter showed progressive reduction with the increasing duration of cryptprchidism. The volume density measurements showed that the seminiferous tubule lumen is increased after cryptorchidism. Degenaration of germ cells resulted in the increase in the tubule lumen (4,6). We showed tubular ectasis increased after surgery and this subject was more prominent in bilateral groups compared with unilateral groups probably due to rapid degeneration of differentiated germ cells in bilateral groups. Many investigators showed significant decreased diameters of seminiferous tubules and thickness of germinal epithelium in cryptorchid testis (23).
During spermatogenesis, the spermatogonial stem cells (SSC) which undergo self-renewal throughout adult life of the animal and produces progeny cells, undergo the complicated differentiation steps. As in other rare stem cell populations, enrichment strategies are required to study SSC biology. It has been demonstrated that during testis development, the total number of SSC per testis increases 39-fold from newborn to adult (24). The number of SSC also appears to increase during the regeneration of spermatogenesis following cytotoxic damage (25, 26). Either testis hyperthermia or cryptorchidism may be used to increase the percentage of SSC invivo. Although these experimental conditions cause depletion of more differentiated germ cells, they do not appear to affect SSC survival and biological activity, thus resulting in an increase in the ratio of SSC vs. the total number of germ cells (27). An effect of heat on spermatogonia in mice had already been reported by Reid, who performed studies on the repoplution of seminiferous tubules after irridiation (28). Differentiation of spermatogonia has also been reported to be arrested in cryptorchid mice (29) and heating the testes of sheep caused a reduction in the production of A1 spermatogonia 20 days later, with no effect on the numbers of A0 spermatogonia (30). Miralgia and Hayashi (31) reported a decrease in the percentage of tubules with spermatogonia from 100 to 89% at day 15 after heating the testes of rats to 43ºC for 30 min. We showed spermatocytes and spermatids are major cells which were affected also spermatogonial number increased after 2 week postoperation and this increase is noticeable in bilateral group compared with unilateral group. In addition, experimentally induced bilateral cryptorchidism provides a useful model for enrichment of spermatogonial stem cells.
According to previous study, experimentally induced unilateral cryptorchidism provides a useful model for study of the role of local factors regulating testis cell apoptosis this suggested that apoptotic cell death induced by bilateral cryptorchidism might be affected by changes in systemic factors. The increase of apoptosis in male germ cell after unilateral cryptorchidism is presumably regulated by local testicular factors (32). Also in this research, the differences indicated between unilateral and bilateral groups maybe arise from this subject.
We believe that the present model is a suitable tool for enrichment of spermatogonial stem cells; also it is useful for treatment of cryptorchidism and further biological research in spermatogenesis.
Acknowledgments
This work was supported by a research grant from Tarbiat Modares University.
We highly thank Shahram Pour Beiranvand and Saeedeh Ebrahimi (Anatomical Sciences Department, Faculty of Medical Sciences, Tarbiat Modares University) for their technical assistant.
Type of Study: Original Article |
References
1. Rockett JC, Map FL, Gares JB. Effects of hyperthermia on spermatogenesis, apoptosis, gene expression and fertility in adult male mice. Biol Reprod 2001; 65: 229-239 [DOI:10.1095/biolreprod65.1.229]
2. Jackson MB. The epidemiology of cryptorchidism. John Radcliffe Hospital Cryptorchidism Research Group. Horm Res 1998; 30: 153-6. [DOI:10.1159/000181052]
3. Ito K, Tanemura K, Gotch H, Kurohmura M, Hayashi Y. Apoptosis-like cell death in experimentally- induced cryptorchidism in adult mice. J Vet Med Sci 1997; 59: 353-359. [DOI:10.1292/jvms.59.353]
4. Kerr JB, Rich KA, de Krester DM. Effects of experimental cryptorchidism on the ultrastructure and function of the sertoli cell and peritubular tissue of the rat testis. Biol Reprod 1979; 21: 823-838. [DOI:10.1095/biolreprod21.4.823]
5. Damber JE, Bergh A, Janson PO. Testicular blood flow and testosterone concentrations in the spermatic venous blood in rats with experimental cryptorchidism. Acta Endocrinol 1978; 88: 611-618. [DOI:10.1530/acta.0.0880611]
6. Kerr JB, Rich KA, de Krester DM. Alteration of the fine structure and androgen secretion of the interstitial cells in the experimentally cryptorchid rat testis. Biol Reprod 1979; 20: 409-422. [DOI:10.1095/biolreprod20.3.409]
7. Hall PF, Kew D, Mita M. The influence of temperature on the function of cultured sertoli cells. Endocrinology 1985; 116: 1926-1932. [DOI:10.1210/endo-116-5-1926]
8. Wang ZQ, Todani T, Watanabe Y, Toki A, Ogura K, Miamoto O, et al. Germ cell degeneration in experimental unilateral cryptorchidism; role of apoptosis. Pediatr Surg Int 1998; 14: 9-13. [DOI:10.1007/s003830050424]
9. Blackshaw AW, Massey PF. Early histological and histochemical changes in the heated and cryptorchid rat testis. Aust J Biol Sci 1978; 31: 53-64.
10. Mieusset R, Bujan L. Testicular heating and its possible contributions to male infertility: a review. Int J Androl 1995; 18: 169-184. [DOI:10.1111/j.1365-2605.1995.tb00408.x]
11. de Rooij DJ, Grootegoud JA. Spermatogonial stem cells. Curr Opin Cell Biol 1998; 10:694-701. [DOI:10.1016/S0955-0674(98)80109-9]
12. Nishimune Y, Aizawa S, Kumatso T. Testicular germ cell differentiation in vivo. Fertil Steril 1978; 29: 95-102. [DOI:10.1016/S0015-0282(16)43045-1]
13. Taki TM, Nickerson PA. Validation of cell volume determination for stereological studies of adrenocortical cells: comparison of values from semifine sections with those of dissociated zona faciculata cells. Am J Anat 1984; 171: 415- 426. [DOI:10.1002/aja.1001710406]
14. World Health Organization 1999 WHO laboratory manual for the examination of human semen and sperm-cervical mucus interaction. 4th ed. Cambridge, UK: Cambridge University Press.
15. Nelson WO. Mammalian spermatogenesis effects of experimental cryptorchidism in the rat and non-descend of the testis in man. Rec Prog Horm Res 1995; 6: 29-62.
16. Yavetz H, Harash B, Paz G, Yogev A, Jaffa A, Lessing JB. Cryptorchidism: incidence and sperm quality in infertile men. Andrologia 1992; 24: 293-297. [DOI:10.1111/j.1439-0272.1992.tb02655.x]
17. Bartlett JMS, Sharp RM. Effect of local heating of the rat testis on the levels in interstitial fluid of a putative paracrine regulator of the leydig cells and its relationship to changes in sertoli cell secretory function. J Reprod Fertil 1987; 80:279-287. [DOI:10.1530/jrf.0.0800279]
18. Chowdhury AK, Stainberger E. A quantitative study of the effect of heat on the germinal epithelium of rat testes. American J Anatomy1964; 115: 509-524. [DOI:10.1002/aja.1001150307]
19. Chowdhury AK, Stainberger E. Early changes in the germinal epithelium of rat testes following exposure to heat. J Reprod Fertil 1970; 22: 205-212. [DOI:10.1530/jrf.0.0220205]
20. Collis PM, Lacy D. Studies on the structure and function of the mammalian testes. Cytological and histochemical observations pn the testis of the rat after a single exposure to heat applied for different lengths of time. Proceedings of the Royal Society of London B 1969; 172: 17-38. [DOI:10.1098/rspb.1969.0009]
21. Thoshihiko S, Hakan B, Aaron JW. Experimentally induced cryptorchidism increases apoptosis in rat testis. Biol Reprod 1994; 51: 865-872. [DOI:10.1095/biolreprod51.5.865]
22. Longquan R, Mohamed S, Marik O. Effects of experimental cryptorchidism on sperm motility and testicular endocrinology in adult male rats. J of Report Develop 2006; 52: 219-228. [DOI:10.1262/jrd.17073]
23. Patkowski JR, Czernik J, Jelen M. The natural course of cryptorchidism in rats and the efficacy of orchidopexy or orchidectomy in its treatment before and after puberty. J Pediater Surg 1992; 27:870-879. [DOI:10.1016/0022-3468(92)90387-M]
24. Shinohara T, Mary R, Avarbock, Brinster RF. Functional analysis of spermatogonial stem cells in steel and cryotorchid infertile mouse model. Dev Biol 2000; 220: 401-411. [DOI:10.1006/dbio.2000.9655]
25. Ogawa t, Ohmura M, Yumura y, Savada H, Honji T, Kubuta Y. Expansion of murine spermatogonial stem cell throught serial transplantation. Biol Reprod 2003; 68; 316-322. [DOI:10.1095/biolreprod.102.004549]
26. Anjamrooz SH, Movahedin M, Mowla SJ, Pour Bairanvand S. Assessment of morphological and functional changes in the mouse testis and epididymal sperms following busulfan treatment. Iran Biomed J 2006; 11; 15-22.
27. Shinohara T, Orwig KE, Avarbock MR, Brinster RL. Remodeling of the post natal mouse testis in accompanied by dramatic changes in stem cell number and niche accessibility. Proc Natl Acad Sci USA 2001; 98: 6186-6191. [DOI:10.1073/pnas.111158198]
28. Ried BO, Mason KA, Withers HR, West J. Effects of hyperthermia and radiation on mouse testis stem cells. Cancer Research 1981; 41: 4453-4457.
29. de Rooij DJ, Okabe M, Nishimune Y. Arrest of spermatogonial differentiation in jsd/jsd, S117H/S117H and cryptorchid mice. Biol Reprod 1999; 61: 842-84. [DOI:10.1095/biolreprod61.3.842]
30. Hochereau-de Reviers MT, Locatelli A, Perreau C, Pisselet C, Setchell BP. Effects of a single brief period of moderate heating of the testes on seminiferous tubules in hypophysectomized rams treated with pituatory extract. J Reprod Fertil 1993; 97: 381-387. [DOI:10.1530/jrf.0.0970381]
31. Miralgia SM, Hayashi H. Histomorphometry of immature rat testis after heating. J Morpho 1993; 217: 65-74. [DOI:10.1002/jmor.1052170106]
32. Shikone T, Billing H, Hsueh AJW. Experimentally induced cryptorchidism increases apoptosis in rat testis. Biol of Reprod 1994; 51: 895-872. [DOI:10.1095/biolreprod51.5.865]
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
