
International Journal of
Reproductive Biomedicine

![]()
![]()
![]()
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
URL: http://ijrm.ir/article-1-3-en.html


Introduction
The surface of endometrium undergo cyclic changes which influence on embryo attachment and implantation. The high level of hormones which produce during ovarian hyperstimulation may effect on the fine structure of endometrium. Several markers of implantation window have been proposed including endometrial morphology, expression and secretion of some cytokine such as leukemia inhibitory factor and appearance of cytoplasmic microprojections, named pinopodes (Lessey, 2001; Lessey, 2001b).These endometrial cell surface projections involve in the pinocytosis of uterine secretion and macromolecules in mice (Nilsson, 1958). The pinopodes have been observed in short time about 24-48 hours at the time of implantation; therefore, their appearance is an indicator of implantation window in mammals (Nikas et al., 1995). Presence and development of pinopodes mainly depend on the ovarian hormones, especially progesterone(Singh et al., 1996). The high concentration of estrogen interferes with normal formation of pinopodes (Martel et al., 1991; Simon et al., 1995). Administration of exogenouse gonadotropic hormones such as hMG and hCG lead to increased secretion of estrogen and progestrone (Kramer and De Wet, 1994). Some investigators showed that ovarian stimulation did not have harmful effect on the endometrial receptivity (Paulson et al., 1997; Kolb et al., 1997). Kolb et al. (1997) reported that an advanced development of the ultrastructure of endometrium surface epithelium in ovarian hyper-stimulated cycles, which may result in an alteretron in the window of implantation. In contrast, there are some reports on a high incidence of dysfunction of endometrium under high physiological level of estrogen and progestrone. The high level of these hormones could affect the endometrial receptivity (Kramer and De Wet, 1994; Karmer et al., 1993; Valbuena et al., 1999 ). Some works have been focused on the ultrastructural characterestics of endometrium surface after controlled ovarian hyperstimulation (COH) using SEM study (Psychoyos and Nikas, 1994; Nikas et al., 1999 ). The objective of this study was to evaluate the correlation between ultrastructural studies for pinopodes expression on the mouse endometrium at the implantation periods after hMG/hCG and progesterone administration.

Materials and Methods
Animals
Forty female NMRI mice aged 6-10 weeks werehoused under 12h light: 12h dark condition. These were randomly divided into two groups:
Group 1: Hyperstimulated mice. Female mice were superovulated with an intraperitoneal injection of 10 i.u. hMG, which were followed 48h later by another injection of 10 i.u. hCG. After superovulation, daily subcutaneous injections of progestrone (1mg/mouse) were performed (Miller, 1979). On the evening of the second injection the mice were rendered pseudopregnant by mating artificially using a plastic sswap, which was turned on the vaginal orifice several times.
Group 2: Control group. Femlae mice were rendered pseudopregnant by mating artificially such as hyperstimulated group.
Tissue preparation
The mice were sacrificed by cervical dislocation at the third and forth days after hCG injection or mating. The samples were obtained from the 1/3 middle part of uterine horns to be fixed immediatly for electrone microscopy studies.
Transmission electron microscopy
Tissues were fixed in 2.5% glutaraldehyde in sodium cacodylate buffer (PH 7.2) at 4˚c for 1.5 h and post fixed in 1% osmium tetroxide for 1 h. The tissues were dehydrated in graded ethanol concentration follwed by acetone then were embedded in epon 812 resin. Semi-thin sections were stained with 1% toluidine blue. Ultra-thin sections were stained with alcoholic uranyl acetate and aqueous lead citrate. The sections were examined using Zeiss transmission electron microscope.
Scanning electron microscopy
The lumen of uterine horns were washed several times by flushing with phosphate buffer to remove mucous secretions, washed with 0.2 M sodium cacodylate buffer (PH 7.2). The samples were fixed primary with 2.5% glutaraldehyd in cacodylate buffer (PH 7.2), after washing with the same buffer, post fixed using 1% osmium tetroxide. The specimens were dehydrated first in unguarded concentrations of ethanol (50%, 70%, 80%, 90% and 100%) and then in acetone. They were dried, mounted and coated with gold particles and examined using a Philips scanning electron microscope.
Results
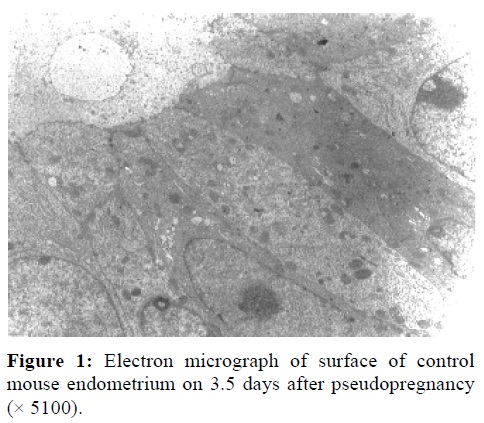
TEMMicrographs of non-stimulated samples on 3.5 days of pseudopregnancy showed that epithelial linning of uterine horns were oval with uchromatin nuclei surrounded by thin marginal heterochromatin at the implantation period. These epithelial cells exhibited some large microvilli (Figure 1). Their cytoplasm was rich in organells such as rough endoplasmic reticulum, polyribosomes and mitochondria. A few lipid droplets with moderate electron density were seen on the subnuclear region.
The organelles of the samples from hyperstimulated and Progestrone injected animals were the same as control group. The stromal cells had polyhedral morphology with elongated nucleus. The intercelluler spaces were smaller than control pregnant groups. The characteristics of stromal cells were protein secreting cells.
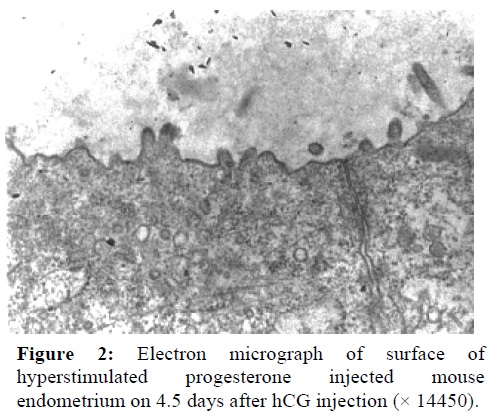
In all groups, the normal microvilli were seen, but these structures disappeared, although some smooth and swollen projections were abundant 3.5 days after hyperstimulation and progesterone injection (Figure 2). It seems that in this group after 4.5 days of hCG injection, microvilli reformed on the other hand the life span of smooth projections was short time (Figure 3).
SEM
Three days after artificial mating the apical cell surface was dome shape and the border of the cells were well defined. The microvilli of these cells were short and the tips of these microvilli dilated. In some cells, the microvilli were disappeared and transformed to fungi shape projection named as pinopodes. These pinopodes were located on the corner of the cells and the remaining surface was smooth (Figure 3). However, on the 4th day, all apical cell surface expressed these projections and a lot of them were collapsed.
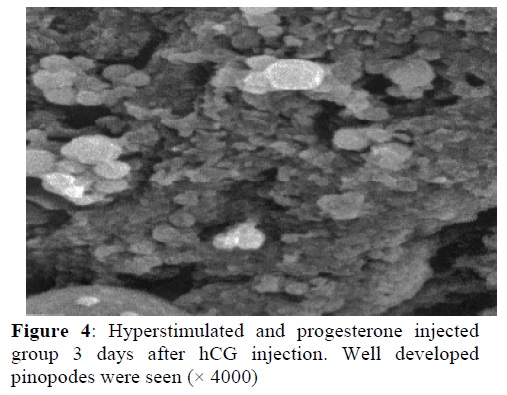
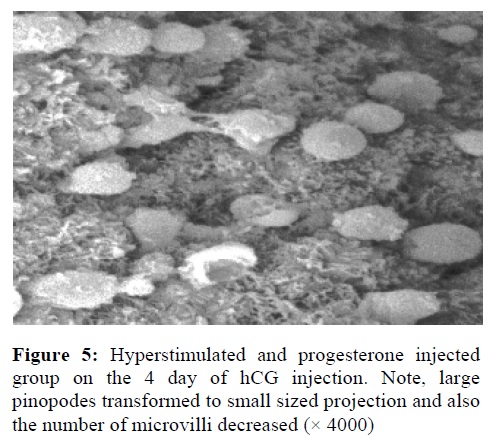
In the hyperstimulated and progesterone injected groups, 3.5 days after hCG the microvilli were disappeared in half of lining cells relatively. Pinopodes were formed and well developed as large spongy projections (Figure 4). In this group, the distinctive morphological changes were observed at the implantation time. A lot of microvilli disappeared and large pinopodes transformed to small projections (Figure 5).
Discussion
Our ultrastructural studies showed that in comparison of apical membrane of all groups, the microvilli were present; although the number of these structures had some changes. However, on the hyperstimulated and progesterone injected groups, the microvilli disappeared and another projections were seen without any glycocalyx on their surfaces on 3.5 day after induction. Our SEM studies showed that these projections were typical pinopodes or fungiform projections, which are implantation marker. It seems that in this group, the microvilli decreased in number and length by fusing together. The progesterone may cause premature expression of this structure.Pinopodes were thought useful marker for endometrium receptivity in many species.Also, our results showed that after administration of progesterone, the well organized pinopodes were expressed over the surface of mouse endometrium. However, these projections were collapsed and transformed to small projections on forth days of hCG and progesterone injection. These characteristics are not apoptotic cells because our TEM study showed that these cells had normal fine morphology. Thus, these observations showed that progesterone injection after hyperstimulation could cause premature expression of pinopodes on the pre implantation time. Our hypothesis is that the presence of pinopodes depends on the progesterone and its balance with estrogen concentration. Also, Stavreus-Evers et al. (2001) showed that pinopod formation in human endometrium is associated with the concentration of progesterone and progesterone receptors.
In contrast to our results, it was shown that under hormone replacement therapy in human, the timing of the nidation window seems to be postponed for some two days and showed that administration of the progesterone antagonist on day 1 of rat pregnancy displaces the time of appearance of fully developed pinopodes from day 5 to 6 or 7 (Arianmanesh et al., 2002). Also, experience from oocyte donation programs in human show that women treated with estradiol and progesterone in hormone-controlled cycles have higher chance of conception when compared with women undergoing ovarian stimulation (Paulson et al., 1990). This may be due to a better priming of endometrium avoiding high oestradiol concentrations (Stavreus-Everse et al., 2001).
Althogh, there are many differences between mouse and human during implantation and pinopodes function,
our results showed that the life span of pinopodes was limited to a short time, which is changed during hyperstimulation regimes. Thus, this alteration in expression of pinopodes results in an alteration in the window of implantation. The alteration in implantation rate following ovarian hyperstimulation in animals may be due to these time changes, and progesterone injection may be effective to premature presence of pinopodes.
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
