
International Journal of
Reproductive Biomedicine
Tue, Aug 4, 2026
[Archive]
Volume 24, Issue 4 (April 2026)
IJRM 2026, 24(4): 349-358 |
Back to browse issues page



Ethics code: IR.UMA.REC.1400.029
![]()
![]()
![]()
Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Haghighat K, Mahmoudi F, Khazali H. Effects of chrysin on Kisspeptin, neurokinin B, and dynorphin neurons and gonadotropin-releasing hormone gene expression in stress-induced rats: An experimental study. IJRM 2026; 24 (4) :349-358
URL: http://ijrm.ir/article-1-3643-en.html
URL: http://ijrm.ir/article-1-3643-en.html



1- Department of Biology, Faculty of Sciences, University of Mohaghegh Ardabili, Ardabil, Iran.
2- Department of Biology, Faculty of Sciences, University of Mohaghegh Ardabili, Ardabil, Iran. ,f.mahmoudi@uma.ac.ir
3- Department of Animal Sciences and Marine Biology, Faculty of Life Sciences and Biotechnology, Shahid Beheshti University, Tehran, Iran.
2- Department of Biology, Faculty of Sciences, University of Mohaghegh Ardabili, Ardabil, Iran. ,
3- Department of Animal Sciences and Marine Biology, Faculty of Life Sciences and Biotechnology, Shahid Beheshti University, Tehran, Iran.
Full-Text [PDF 926 kb]
(293 Downloads)
| Abstract (HTML) (366 Views)
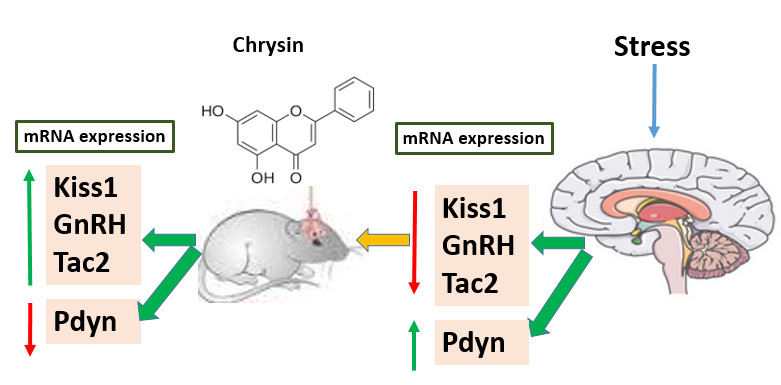
Full-Text: (49 Views)
1. Introduction
Several signaling systems are converged in the hypothalamus to regulate the reproductive process (1). Physical or psychological stress is accompanied by the activation of the hypothalamic-pituitary-adrenal (HPA) axis, which in turn affects all aspects of endocrine systems such as the hypothalamus-pituitary-gonadal (HPG) axis. In fact, stress inhibits the release of gonadotropin-releasing hormone (GnRH) and luteinizing hormone (LH), and chronic stress may result in infertility (2, 3).
Kisspeptin, encoded by Kiss1 gene, is expressed in the hypothalamus, especially arcuate nucleus (ARC), to regulate tonic GnRH/LH secretion (4, 5). Kiss1 is involved in the direct stimulation of GnRH release due to expression of its receptor named G-Kiss1 coupled receptor-54 on GnRH neurons (4, 6). The stimulatory effects of Kiss1 are completely established on the HPG axis (6-8). Recent studies introduce Kiss1 as a potential therapeutic target for management of infertility and reproductive disorders. Involvement of Kisspeptin, neurokinin B, and dynorphin (KNDy) neurons has been demonstrated in stress suppression of the HPG axis, and stress causes significant decrease in GnRH and kisspeptin signalling pathways (9-11).
Neurokinin B (NKB)encoded by Tachykinin 2 (Tac2) gene, is expressed predominantly in the hypothalamus in ARC and exerts its physiological effects via binding neurokinin 3 receptor (12, 13). Kisspeptin neurons (named KNDy neurons) of ARC co-express NKB and dynorphin (Dyn) to regulate GnRH pulse generator and negative feedback controlling of testosterone on HPG axis (12). In fact, neurokinin 3 receptor is expressed on kisspeptin and GnRH neurons, and NKB acts upstream of kisspeptin to regulate GnRH/LH release via direct and indirect signalling pathways (12, 14).
Dyn encoded by Pdyn gene, is an endogenous opioid peptide that acts via binding κ-opioid receptor (KOR). The Dyn/KOR system suppresses GnRH/LH release (14). Also, Dyn is a key factor in mediating stress. Anxiety increases the HPA axis activity and results in high release of corticotropin-releasing hormone (CRH) and Dyn. High levels of Dyn, in turn, take part in the mediation of stress-induced responses (15).
Chrysin is mainly isolated from honey, propolis, and several herbal medicines such as Passiflora incarnata, Passiflora coerulea, and Oroxylum indicum (16, 17). Some studies indicated that chrysin can protect testicular tissue against chemical toxicants (18, 19). However, some studies reported that honey and herbal medicines rich in chrysin did not alter testosterone concentration in males. Previous in vitro studies suggested that aromatase inhibitory effect of chrysin may be involved in its testosterone boosting effect (20, 21). Also, some studies showed that increasing the cAMP-induced steroidogenic acute regulatory gene expression or stimulating LH secretion may be involved in the increasing effect of chrysin on testosterone (22, 23). Also, it has been indicated that chrysin may inhibit stressful behaviors via affecting the function of gamma-aminobutyric acid-A (GABAA) receptors (24).
Since the molecular mechanisms underlying the stimulatory effects of chrysin on testosterone secretion and HPG axis are unclear. The present study aimed to determine the effects of chrysin on gene expression of Kiss1, Tac2, and Pdyn upstream GnRH neurons in intact and stress-subjected rats.
2. Materials and Methods
2.1. Materials
The rats were supplied by the Pasteur Institute, Tehran, Iran. Chrysin (Cas No. 480-40-0, Co. USA), cDNA kit (Biotech rabbit, Germany), and SYBR Green kit (Takara Bio Inc., Japan) were purchased from Sigma Aldrich, U.S.A.
2.2. Animal
In this experimental study, 30 male Wistar rats (200 ± 10 gr, 8 wk) (n = 5/each) were used. The rats were housed in an air-conditioned room with a temperature of 22 ± 2°C, a humidity of 50 ± 10%, and a 12-hr light/dark cycle for 7 days in order to allow the rats to acclimate to their new environment. The work was conducted in the Research Physiology Laboratory, University of Mohaghegh Ardabili, Ardabil, Iran, from April 2024 to August 2024.
2.3. Sample size
Unlike clinical and descriptive studies that require large sample sizes, this experimental study follows animal research protocols, which limit the number of subjects to a maximum of 5, except in behavioral studies, where the sample size may be increased to 7.
2.4. Surgical procedure
First, an intraperitoneal injection of xylazine (10 mg/kg) and ketamine (80 mg/kg) was used to put the animal to sleep. Then the position of the animal was adjusted in stereotaxy. Based on the Paxinos and Watson Atlas coordinates (AP = 0.84 mm, ML = 00, DV = 6.5 mm), the cannula was placed in the skull. Following a week of recuperation in the laboratory, the animals were given drugs (third cerebral ventricle) by using a Hamilton syringe that was fastened to a polyethylene tube 20.
2.5. Acute stress induction and treatment
First, the animals were randomly grouped (n = 5). The injection was done to each group as follows: group I: received saline as a control group. Group II: received saline as a stress group. Groups III and IV: received Chrysin (40 or 20 μg, intracerebroventricular injection [ICV], single dose), as a stress-induced group. Groups V and VI: received Chrysin (40 or 20 μg, ICV, single dose), as an intact group (did not undergo stress exposure) (25). For acute stress induction, 30 min after receiving chrysin, rats were placed in a restraining box (length 18 cm, diameter 5 cm). Then, the boxes were moved to a quiet, dark room for 2 hr. All injections were performed between 9 and 11 AM.
2.6. Reverse transcriptase polymerase chain reaction
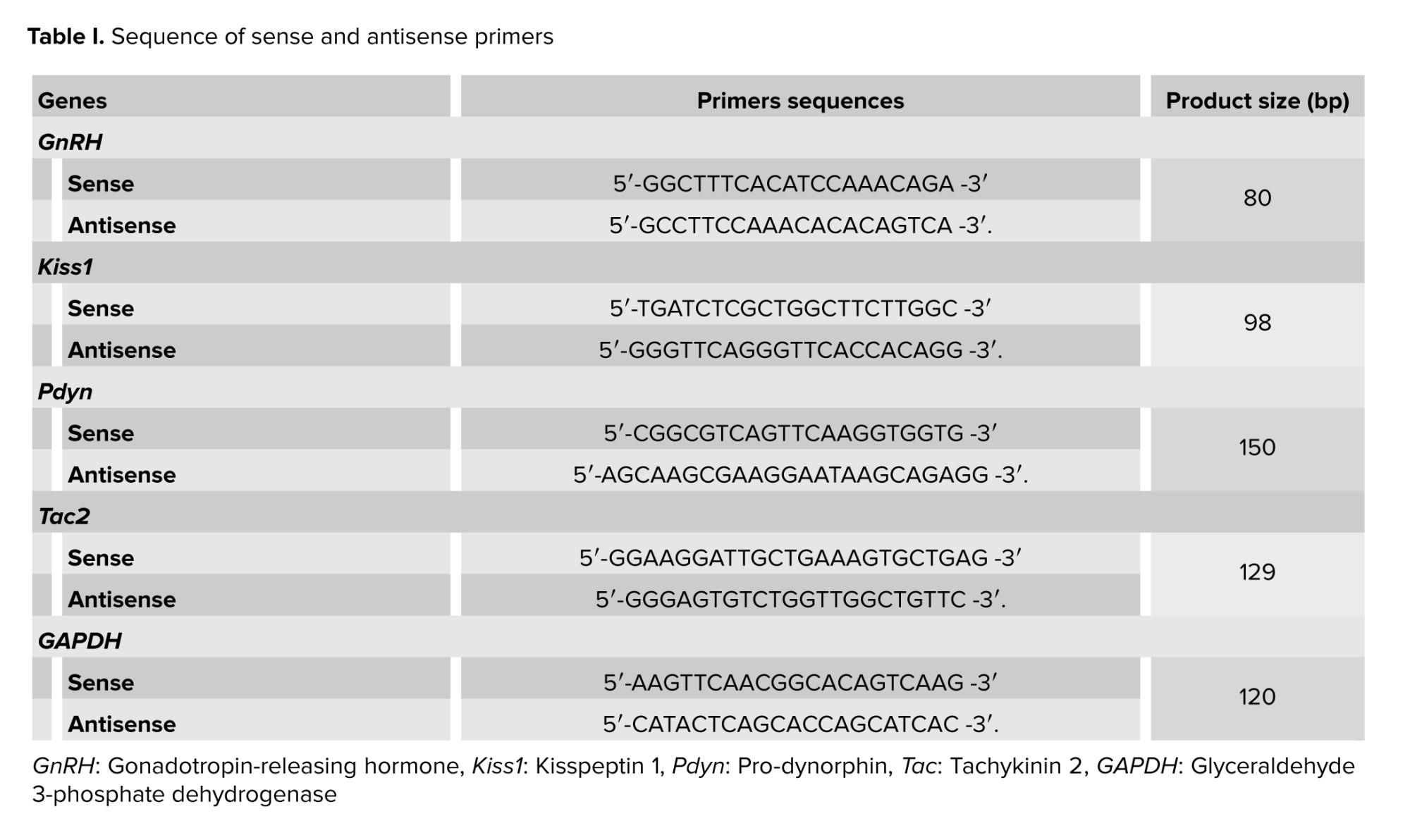
The hypothalamus tissues were removed from the groups (n = 5/each) and stored at -80°C. The total RNA was extracted from the hypothalamic samples utilizing the TRIzol reagent according to the kit instructions. The RNA concentration was determined using a Nano Drop device. Then, a cDNA synthesis kit was used to convert RNA to cDNA. To perform real-time polymerase chain reaction, SYBR Green kit was used according to the manufacturer’s directions. The conditions for thermal cycling were following: 1 cycle for 15 min at 95°C was carried out. After, it went through 40 cycles, which included 20 sec of denaturation at 95°C, 15 sec of annealing at 60°C, and 10 sec of extension at 72°C (5). The relative expression of GnRH, Kiss1, Pdyn and Tac2 were normalized to GAPDH and relative changes in gene expression were calculated by the 2_ΔΔCT. Specific oligo nucleotide sequences for forward and reverse primers are listed in table I.
2.7. Ethical Considerations
This experimental study was approved by the Research Ethics Committee of the University of Mohaghegh Ardabili, Ardabil, Iran (Code: IR.UMA.REC.1400.029). In order to observe ethical considerations and prevent animal suffering, all animals were anesthetized before performing surgical procedures.
2.8. Statistical Analysis
The data were analyzed using SPSS software (version 16) and one-way ANOVA. The next step was Tukey's post hoc test. The criterion of statistical significance was a p ≤ 0.05. Mean ± SEM was used to express the obtained results.
3. Results
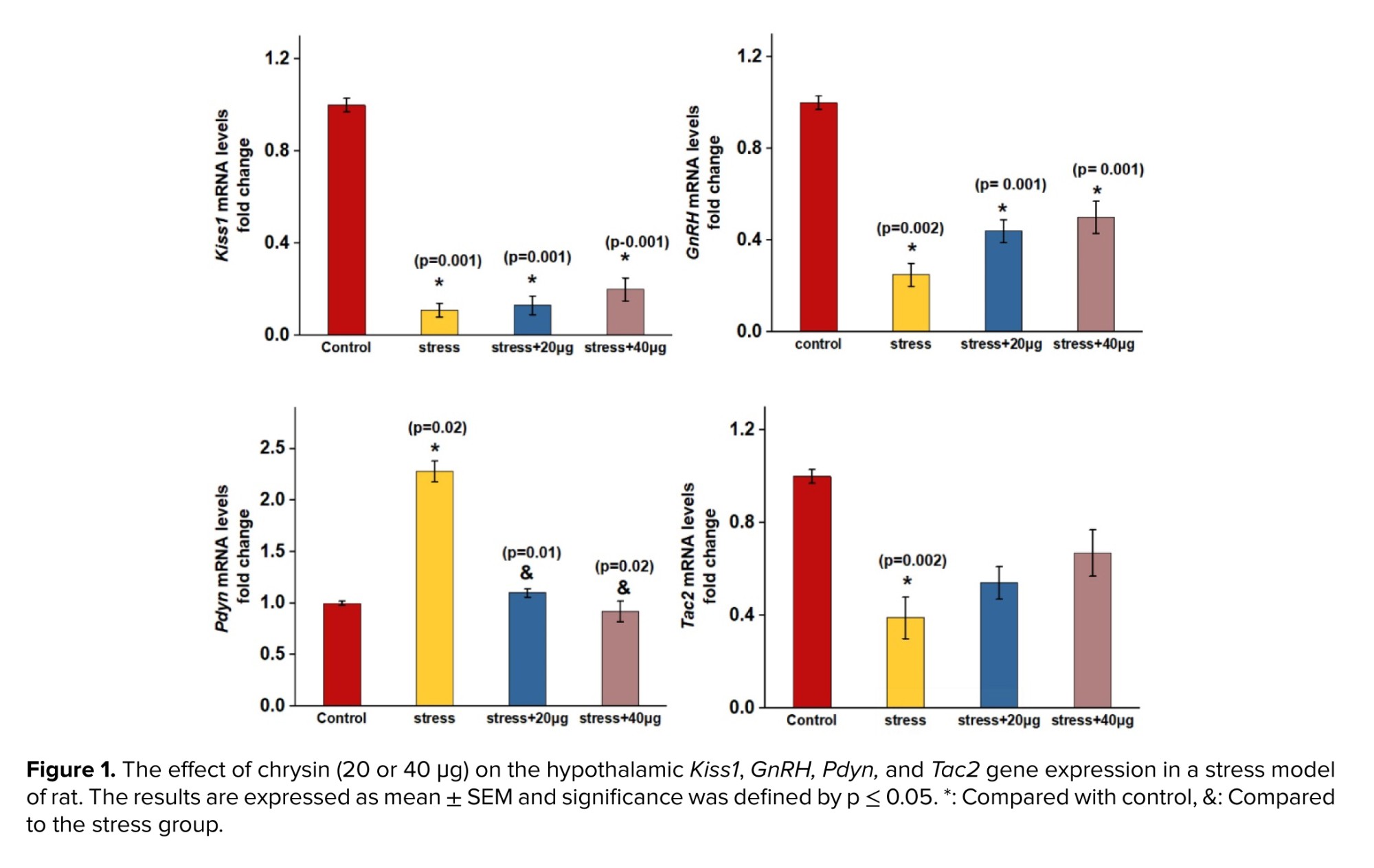
In figure 1, expression of Kiss1, GnRH, and Tac2 genes were downregulated in the stress group, compared to the control group. The decrease was statistically significant. However, no significant increase was observed in the expression of Kiss, GnRH, and Tac2 genes in rats treated with chrysin )40 or 20 μg (compared to the stress group. Also, the expression of Kiss1, GnRH, and Tac2 decreased in the group receiving chrysin (40 or 20 μg (compared to the control group, which was a significant decrease in the expression of Kiss1 and GnRH genes. Moreover, the expression of Pdyn in the stress group increased significantly compared to control group. Chrysin (40 or 20 μg) injection caused a significant decrease in the expression of the Pdyn gene relative to the stress group. No significant change was observed in Pdyn gene expression in groups receiving chrysin (40 or 20 μg (compared to the control group.
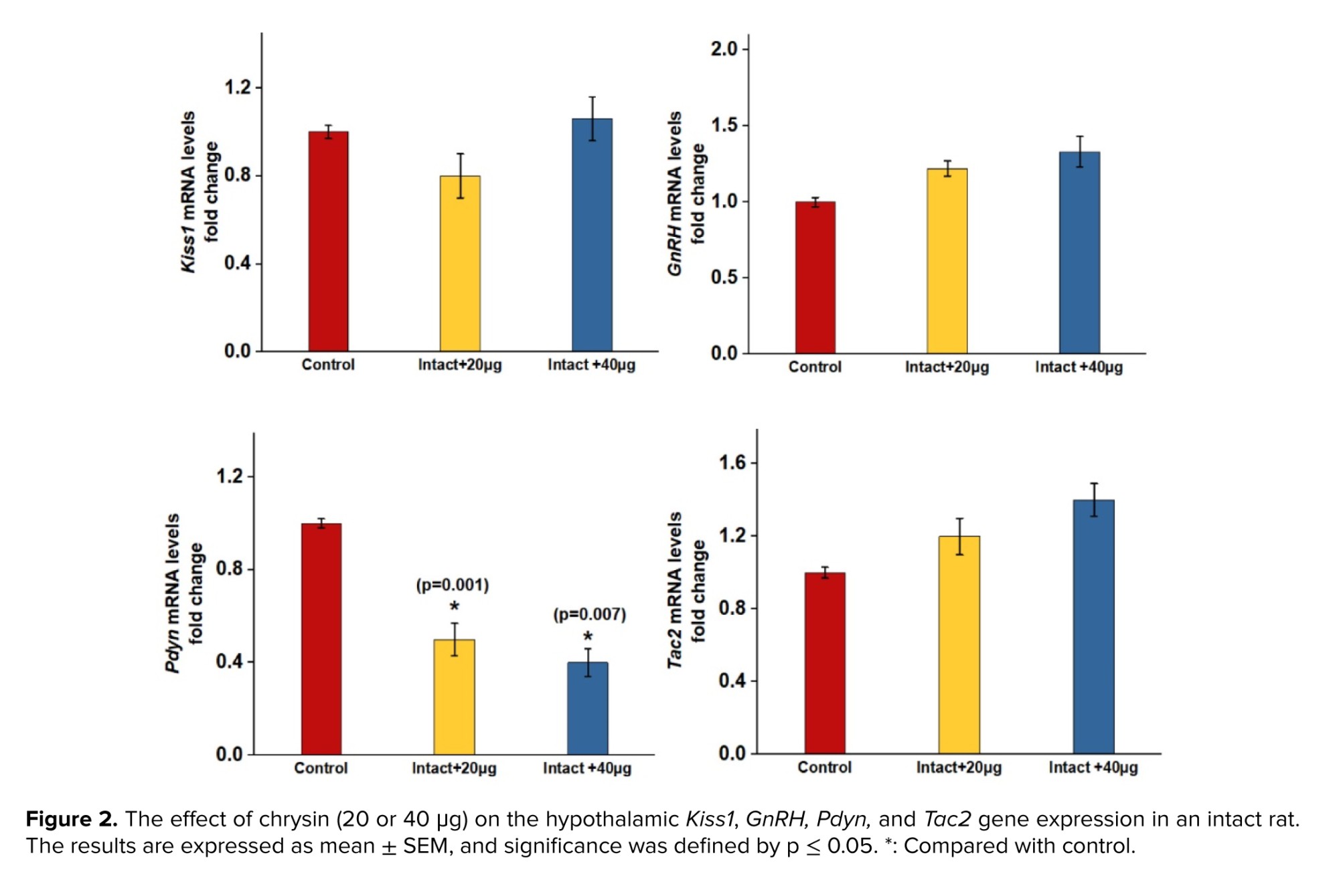
As shown in figure 2, no significant difference was observed in the expression of Kiss, GnRH, and Tac2 in the intact group receiving chrysin (40 or 20 μg) compared to the control group. However, the expression of Pdyn in rats treated with chrysin (40 or 20 μg) decreased significantly compared to the control group.
4. Discussion
The present results indicated that ICV of chrysin did not significantly alter GnRH, Kiss1, and Tac2 gene expression in intact rats compared to control group. However, the expression of Pdyn gene significantly declined in intact rats receiving chrysin compared to control group. Chrysin’s testosterone increasing effect was indicated in some previous studies (18). However, in some studies, stimulatory effects of chrysin, honey, and passion flower extract (both rich sources of chrysin) were not demonstrated in healthy young men (20, 26). KNDy take part in controlling the GnRH/LH release and testosterone secretion (1). Based on several previous studies, kisspeptin and NKB act as stimulatory GnRH neurons, while Dyn inhibits GnRH/LH release directly or indirectly via downregulating kisspeptin and NKB signalling pathway (1, 4, 27). Since the mechanisms by which chrysin influences reproduction are not clear, the present study was conducted to investigate some intra-hypothalamic possible mechanisms of the increasing effect of chrysin on testosterone.
A study on both young and old men showed that treatment with passion flower extract, a rich source of chrysin, did not affect testosterone concentration and libido in younger men; however, it caused an increase in libido and testosterone concentration in older men (26). It is reported that passion flower extract may be effective when delivered to subjects with low baseline testosterone (26, 28). Low testosterone is usually associated with aging, stress, diabetes, and other disorders (27). In the second part of the study, the effects of ICV of chrysin were investigated on hypothalamic Kiss1, Tac2, and Pdyn expression gene in stress-subjected rats, a disorder which is associated with low baseline testosterone levels.
The results indicated that induction of stress significantly decreased the hypothalamic expression of GnRH and Kiss1 genes. The present results are in agreement with several previous studies, which showed that restraint or immobilization stress, activation of HPA axis, and CRH injection suppresses GnRH, gonadotropin, steroid hormones secretion, and kisspeptin synthesis (9-11).
The present data shows that restraint stress did not significantly decline hypothalamic expression of Tac2. These findings are in line with the study, which demonstrated that acute restraint stress did not alter Tac2 gene expression in the hypothalamus of mice (11).
Restraint stress caused a significant increase in Pdyn expression gene in comparison to control group. These findings are consistent with several previous studies, which indicated the involvement of Dyn-KOR system in mediating anxiogenic behaviors. Dny and κ opioid receptors are highly expressed in brain areas that are associated with stress responses, such as hypothalamic paraventricular nucleus (PVN), hippocampus, amygdala, and stress exposure that causes rising of Dny release.
Based on previous studies, the connection between Dyn-KOR and CRH has been established. A population of CRH neurons co-expresses Dny in the hypothalamus. The acute stress activates CRH neurons and κ opioid receptor in the PVN, and CRH causes Dny release. Also, it has been shown that CRH-induced stressful behaviors are suppressed by KOR antagonists (15, 29).
Injection of chrysin significantly decreased the Pdyn expression gene compared to stress group. Since chrysin did not affect GnRH, Kiss1, and Tac2 expression genes in stressed rats, thus the suppressive effects of chrysin on hypothalamic Pdyn expression gene may be consistent with Dny role in mediating stress.
It has been shown that chrysin binds to the GABAA receptors and activates the GABAergic system. The antagonists of GABAA receptor can block chrysin's anti-stress influences (18). A previous study indicated that Dyn-KOR system inhibits presynaptic release of GABA (30). The GABAergic neurons innervate CRH neurons in the PVN of the hypothalamus, and GABA suppresses the activity of HPA axis by binding to GABAA receptors (31). Also, previous studies have showed that chrysin was able to decrease CRH and corticosterone concentration levels in rats submitted to stress (32). So, activation of the hypothalamic GABAergic neurons and inhibition of HPA axis may be a possible mechanism for suppressive effects of chrysin on Pdyn gene expression in stress responses.
5. Conclusion
The present results indicated that the third cerebral ventricular injection of chrysin significantly declined Pdyn gene expression, but it did not significantly alter the hypothalamic GnRH, Kiss1, and Tac2 gene expression in intact rats compared to control group. In stress-subjected rats, chrysin can significantly decrease Pdyn gene expression. However, it did not alter GnRH, Kiss1, and Tac2 gene expression in stressed rats. The effects of chrysin on reproduction may not be mediated via alteration of GnRH and KNDy neurons upstream of GnRH neurons. Downregulation of hypothalamic Dny may be linked to anxiolytic potency of chrysin. The study limited the evaluation of chrysin to measuring gene expression. Protein-level validation is currently lacking and should be included in future investigations.
Data Availability
The data are available from the corresponding author on reasonable request.
Author Contributions
F. Mahmoudi and Kh. Haghighat designed the study and conducted the research. F. Mahmoudi and H. Khazali monitored, evaluated, and analyzed the result of the study. Further, F. Mahmoudi reviewed the article. All authors approved the final manuscript and take responsibility for the integrity of the data.
Acknowledgments
The authors wish to appreciate the University of Mohaghegh Ardabili, Ardabil, Iran for generously providing financial support and supplying the required laboratory and equipment to conduct this study. The present article has been extracted from Ph.D. dissertation (1400/13/5765) and the non-use of artificial intelligence.
Conflict of Interest
The author declares that there is no conflict of interest.
Several signaling systems are converged in the hypothalamus to regulate the reproductive process (1). Physical or psychological stress is accompanied by the activation of the hypothalamic-pituitary-adrenal (HPA) axis, which in turn affects all aspects of endocrine systems such as the hypothalamus-pituitary-gonadal (HPG) axis. In fact, stress inhibits the release of gonadotropin-releasing hormone (GnRH) and luteinizing hormone (LH), and chronic stress may result in infertility (2, 3).
Kisspeptin, encoded by Kiss1 gene, is expressed in the hypothalamus, especially arcuate nucleus (ARC), to regulate tonic GnRH/LH secretion (4, 5). Kiss1 is involved in the direct stimulation of GnRH release due to expression of its receptor named G-Kiss1 coupled receptor-54 on GnRH neurons (4, 6). The stimulatory effects of Kiss1 are completely established on the HPG axis (6-8). Recent studies introduce Kiss1 as a potential therapeutic target for management of infertility and reproductive disorders. Involvement of Kisspeptin, neurokinin B, and dynorphin (KNDy) neurons has been demonstrated in stress suppression of the HPG axis, and stress causes significant decrease in GnRH and kisspeptin signalling pathways (9-11).
Neurokinin B (NKB)encoded by Tachykinin 2 (Tac2) gene, is expressed predominantly in the hypothalamus in ARC and exerts its physiological effects via binding neurokinin 3 receptor (12, 13). Kisspeptin neurons (named KNDy neurons) of ARC co-express NKB and dynorphin (Dyn) to regulate GnRH pulse generator and negative feedback controlling of testosterone on HPG axis (12). In fact, neurokinin 3 receptor is expressed on kisspeptin and GnRH neurons, and NKB acts upstream of kisspeptin to regulate GnRH/LH release via direct and indirect signalling pathways (12, 14).
Dyn encoded by Pdyn gene, is an endogenous opioid peptide that acts via binding κ-opioid receptor (KOR). The Dyn/KOR system suppresses GnRH/LH release (14). Also, Dyn is a key factor in mediating stress. Anxiety increases the HPA axis activity and results in high release of corticotropin-releasing hormone (CRH) and Dyn. High levels of Dyn, in turn, take part in the mediation of stress-induced responses (15).
Chrysin is mainly isolated from honey, propolis, and several herbal medicines such as Passiflora incarnata, Passiflora coerulea, and Oroxylum indicum (16, 17). Some studies indicated that chrysin can protect testicular tissue against chemical toxicants (18, 19). However, some studies reported that honey and herbal medicines rich in chrysin did not alter testosterone concentration in males. Previous in vitro studies suggested that aromatase inhibitory effect of chrysin may be involved in its testosterone boosting effect (20, 21). Also, some studies showed that increasing the cAMP-induced steroidogenic acute regulatory gene expression or stimulating LH secretion may be involved in the increasing effect of chrysin on testosterone (22, 23). Also, it has been indicated that chrysin may inhibit stressful behaviors via affecting the function of gamma-aminobutyric acid-A (GABAA) receptors (24).
Since the molecular mechanisms underlying the stimulatory effects of chrysin on testosterone secretion and HPG axis are unclear. The present study aimed to determine the effects of chrysin on gene expression of Kiss1, Tac2, and Pdyn upstream GnRH neurons in intact and stress-subjected rats.
2. Materials and Methods
2.1. Materials
The rats were supplied by the Pasteur Institute, Tehran, Iran. Chrysin (Cas No. 480-40-0, Co. USA), cDNA kit (Biotech rabbit, Germany), and SYBR Green kit (Takara Bio Inc., Japan) were purchased from Sigma Aldrich, U.S.A.
2.2. Animal
In this experimental study, 30 male Wistar rats (200 ± 10 gr, 8 wk) (n = 5/each) were used. The rats were housed in an air-conditioned room with a temperature of 22 ± 2°C, a humidity of 50 ± 10%, and a 12-hr light/dark cycle for 7 days in order to allow the rats to acclimate to their new environment. The work was conducted in the Research Physiology Laboratory, University of Mohaghegh Ardabili, Ardabil, Iran, from April 2024 to August 2024.
2.3. Sample size
Unlike clinical and descriptive studies that require large sample sizes, this experimental study follows animal research protocols, which limit the number of subjects to a maximum of 5, except in behavioral studies, where the sample size may be increased to 7.
2.4. Surgical procedure
First, an intraperitoneal injection of xylazine (10 mg/kg) and ketamine (80 mg/kg) was used to put the animal to sleep. Then the position of the animal was adjusted in stereotaxy. Based on the Paxinos and Watson Atlas coordinates (AP = 0.84 mm, ML = 00, DV = 6.5 mm), the cannula was placed in the skull. Following a week of recuperation in the laboratory, the animals were given drugs (third cerebral ventricle) by using a Hamilton syringe that was fastened to a polyethylene tube 20.
2.5. Acute stress induction and treatment
First, the animals were randomly grouped (n = 5). The injection was done to each group as follows: group I: received saline as a control group. Group II: received saline as a stress group. Groups III and IV: received Chrysin (40 or 20 μg, intracerebroventricular injection [ICV], single dose), as a stress-induced group. Groups V and VI: received Chrysin (40 or 20 μg, ICV, single dose), as an intact group (did not undergo stress exposure) (25). For acute stress induction, 30 min after receiving chrysin, rats were placed in a restraining box (length 18 cm, diameter 5 cm). Then, the boxes were moved to a quiet, dark room for 2 hr. All injections were performed between 9 and 11 AM.
2.6. Reverse transcriptase polymerase chain reaction
The hypothalamus tissues were removed from the groups (n = 5/each) and stored at -80°C. The total RNA was extracted from the hypothalamic samples utilizing the TRIzol reagent according to the kit instructions. The RNA concentration was determined using a Nano Drop device. Then, a cDNA synthesis kit was used to convert RNA to cDNA. To perform real-time polymerase chain reaction, SYBR Green kit was used according to the manufacturer’s directions. The conditions for thermal cycling were following: 1 cycle for 15 min at 95°C was carried out. After, it went through 40 cycles, which included 20 sec of denaturation at 95°C, 15 sec of annealing at 60°C, and 10 sec of extension at 72°C (5). The relative expression of GnRH, Kiss1, Pdyn and Tac2 were normalized to GAPDH and relative changes in gene expression were calculated by the 2_ΔΔCT. Specific oligo nucleotide sequences for forward and reverse primers are listed in table I.
2.7. Ethical Considerations
This experimental study was approved by the Research Ethics Committee of the University of Mohaghegh Ardabili, Ardabil, Iran (Code: IR.UMA.REC.1400.029). In order to observe ethical considerations and prevent animal suffering, all animals were anesthetized before performing surgical procedures.
2.8. Statistical Analysis
The data were analyzed using SPSS software (version 16) and one-way ANOVA. The next step was Tukey's post hoc test. The criterion of statistical significance was a p ≤ 0.05. Mean ± SEM was used to express the obtained results.
3. Results
In figure 1, expression of Kiss1, GnRH, and Tac2 genes were downregulated in the stress group, compared to the control group. The decrease was statistically significant. However, no significant increase was observed in the expression of Kiss, GnRH, and Tac2 genes in rats treated with chrysin )40 or 20 μg (compared to the stress group. Also, the expression of Kiss1, GnRH, and Tac2 decreased in the group receiving chrysin (40 or 20 μg (compared to the control group, which was a significant decrease in the expression of Kiss1 and GnRH genes. Moreover, the expression of Pdyn in the stress group increased significantly compared to control group. Chrysin (40 or 20 μg) injection caused a significant decrease in the expression of the Pdyn gene relative to the stress group. No significant change was observed in Pdyn gene expression in groups receiving chrysin (40 or 20 μg (compared to the control group.
As shown in figure 2, no significant difference was observed in the expression of Kiss, GnRH, and Tac2 in the intact group receiving chrysin (40 or 20 μg) compared to the control group. However, the expression of Pdyn in rats treated with chrysin (40 or 20 μg) decreased significantly compared to the control group.
4. Discussion
The present results indicated that ICV of chrysin did not significantly alter GnRH, Kiss1, and Tac2 gene expression in intact rats compared to control group. However, the expression of Pdyn gene significantly declined in intact rats receiving chrysin compared to control group. Chrysin’s testosterone increasing effect was indicated in some previous studies (18). However, in some studies, stimulatory effects of chrysin, honey, and passion flower extract (both rich sources of chrysin) were not demonstrated in healthy young men (20, 26). KNDy take part in controlling the GnRH/LH release and testosterone secretion (1). Based on several previous studies, kisspeptin and NKB act as stimulatory GnRH neurons, while Dyn inhibits GnRH/LH release directly or indirectly via downregulating kisspeptin and NKB signalling pathway (1, 4, 27). Since the mechanisms by which chrysin influences reproduction are not clear, the present study was conducted to investigate some intra-hypothalamic possible mechanisms of the increasing effect of chrysin on testosterone.
A study on both young and old men showed that treatment with passion flower extract, a rich source of chrysin, did not affect testosterone concentration and libido in younger men; however, it caused an increase in libido and testosterone concentration in older men (26). It is reported that passion flower extract may be effective when delivered to subjects with low baseline testosterone (26, 28). Low testosterone is usually associated with aging, stress, diabetes, and other disorders (27). In the second part of the study, the effects of ICV of chrysin were investigated on hypothalamic Kiss1, Tac2, and Pdyn expression gene in stress-subjected rats, a disorder which is associated with low baseline testosterone levels.
The results indicated that induction of stress significantly decreased the hypothalamic expression of GnRH and Kiss1 genes. The present results are in agreement with several previous studies, which showed that restraint or immobilization stress, activation of HPA axis, and CRH injection suppresses GnRH, gonadotropin, steroid hormones secretion, and kisspeptin synthesis (9-11).
The present data shows that restraint stress did not significantly decline hypothalamic expression of Tac2. These findings are in line with the study, which demonstrated that acute restraint stress did not alter Tac2 gene expression in the hypothalamus of mice (11).
Restraint stress caused a significant increase in Pdyn expression gene in comparison to control group. These findings are consistent with several previous studies, which indicated the involvement of Dyn-KOR system in mediating anxiogenic behaviors. Dny and κ opioid receptors are highly expressed in brain areas that are associated with stress responses, such as hypothalamic paraventricular nucleus (PVN), hippocampus, amygdala, and stress exposure that causes rising of Dny release.
Based on previous studies, the connection between Dyn-KOR and CRH has been established. A population of CRH neurons co-expresses Dny in the hypothalamus. The acute stress activates CRH neurons and κ opioid receptor in the PVN, and CRH causes Dny release. Also, it has been shown that CRH-induced stressful behaviors are suppressed by KOR antagonists (15, 29).
Injection of chrysin significantly decreased the Pdyn expression gene compared to stress group. Since chrysin did not affect GnRH, Kiss1, and Tac2 expression genes in stressed rats, thus the suppressive effects of chrysin on hypothalamic Pdyn expression gene may be consistent with Dny role in mediating stress.
It has been shown that chrysin binds to the GABAA receptors and activates the GABAergic system. The antagonists of GABAA receptor can block chrysin's anti-stress influences (18). A previous study indicated that Dyn-KOR system inhibits presynaptic release of GABA (30). The GABAergic neurons innervate CRH neurons in the PVN of the hypothalamus, and GABA suppresses the activity of HPA axis by binding to GABAA receptors (31). Also, previous studies have showed that chrysin was able to decrease CRH and corticosterone concentration levels in rats submitted to stress (32). So, activation of the hypothalamic GABAergic neurons and inhibition of HPA axis may be a possible mechanism for suppressive effects of chrysin on Pdyn gene expression in stress responses.
5. Conclusion
The present results indicated that the third cerebral ventricular injection of chrysin significantly declined Pdyn gene expression, but it did not significantly alter the hypothalamic GnRH, Kiss1, and Tac2 gene expression in intact rats compared to control group. In stress-subjected rats, chrysin can significantly decrease Pdyn gene expression. However, it did not alter GnRH, Kiss1, and Tac2 gene expression in stressed rats. The effects of chrysin on reproduction may not be mediated via alteration of GnRH and KNDy neurons upstream of GnRH neurons. Downregulation of hypothalamic Dny may be linked to anxiolytic potency of chrysin. The study limited the evaluation of chrysin to measuring gene expression. Protein-level validation is currently lacking and should be included in future investigations.
Data Availability
The data are available from the corresponding author on reasonable request.
Author Contributions
F. Mahmoudi and Kh. Haghighat designed the study and conducted the research. F. Mahmoudi and H. Khazali monitored, evaluated, and analyzed the result of the study. Further, F. Mahmoudi reviewed the article. All authors approved the final manuscript and take responsibility for the integrity of the data.
Acknowledgments
The authors wish to appreciate the University of Mohaghegh Ardabili, Ardabil, Iran for generously providing financial support and supplying the required laboratory and equipment to conduct this study. The present article has been extracted from Ph.D. dissertation (1400/13/5765) and the non-use of artificial intelligence.
Conflict of Interest
The author declares that there is no conflict of interest.
Type of Study: Original Article |
Subject:
Reproductive Genetics
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
