
International Journal of
Reproductive Biomedicine
Sun, Jul 26, 2026
[Archive]
Volume 24, Issue 3 (March 2026)
IJRM 2026, 24(3): 193-208 |
Back to browse issues page



Ethics code: IR.SBMU.MSP.REC.1403.331
![]()
![]()
![]()
Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Babakhanzadeh E, Khodadadian A, Nazari M, Mozhdeh M, Fazeli J, Vahidi S, et al . A novel hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 regulatory axis as a non-invasive biomarker for predicting sperm retrieval outcomes in non-obstructive azoospermia: A case-control study. IJRM 2026; 24 (3) :193-208
URL: http://ijrm.ir/article-1-3726-en.html
URL: http://ijrm.ir/article-1-3726-en.html
Emad Babakhanzadeh1 

 , Ali Khodadadian2 , Majid Nazari2 , Mahya Mozhdeh3 , Javad Fazeli2 , Serajoddin Vahidi4 , Mohammad Hassan Jafari5 , Mohammadreza Dehghani2 , Soudeh Ghafouri-Fard *6 , Nasrin Ghasemi7
, Ali Khodadadian2 , Majid Nazari2 , Mahya Mozhdeh3 , Javad Fazeli2 , Serajoddin Vahidi4 , Mohammad Hassan Jafari5 , Mohammadreza Dehghani2 , Soudeh Ghafouri-Fard *6 , Nasrin Ghasemi7
, Ali Khodadadian2 , Majid Nazari2 , Mahya Mozhdeh3 , Javad Fazeli2 , Serajoddin Vahidi4 , Mohammad Hassan Jafari5 , Mohammadreza Dehghani2 , Soudeh Ghafouri-Fard *6 , Nasrin Ghasemi7
1- Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran. & Department of Medical Genetics, School of Medicine, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
2- Department of Medical Genetics, School of Medicine, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
3- Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran.
4- Andrology Research Center, Yazd Reproductive Sciences Institute, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
5- Department of Medical Biotechnology, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
6- Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran. ,s.ghafourifard@sbmu.ac.ir
7- Abortion Research Center, Yazd Reproductive Sciences Institute, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
2- Department of Medical Genetics, School of Medicine, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
3- Department of Biology, Science and Research Branch, Islamic Azad University, Tehran, Iran.
4- Andrology Research Center, Yazd Reproductive Sciences Institute, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
5- Department of Medical Biotechnology, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
6- Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran. ,
7- Abortion Research Center, Yazd Reproductive Sciences Institute, Shahid Sadoughi University of Medical Sciences, Yazd, Iran.
Full-Text [PDF 2617 kb]
(369 Downloads)
| Abstract (HTML) (354 Views)
2. Materials and Methods
3. Results
5. Conclusion
Full-Text: (51 Views)
1. Introduction
Male infertility is a significant global health concern, with non-obstructive azoospermia (NOA) representing one of its most severe and challenging forms (1-3). NOA is defined by the complete absence of sperm in the ejaculate despite the lack of any physical obstruction in the reproductive tract. It affects approximately 1% of the general male population and up to 10-15% of infertile men (4). Importantly, the etiology of NOA remains idiopathic in nearly 70% of cases, even after extensive clinical evaluation, highlighting a substantial gap in the molecular understanding of spermatogenic failure (5).
Microsurgical testicular sperm extraction (micro-TESE) has enabled some men with NOA to have their own biological children (6); however, its success rate is limited to roughly 50% (7). Moreover, currently used clinical predictors of sperm retrieval success, such as testicular volume, serum follicle-stimulating hormone (FSH) or inhibin levels, age, and histopathology are inconsistent and unreliable (8). As a result, many individuals undergo an invasive and costly procedure with uncertain outcomes, underscoring the urgent need for reliable, non-invasive biomarkers that can predict micro-TESE success and improve individual counseling and clinical decision-making.
In recent years, non-coding RNAs (ncRNAs), particularly circular RNAs (circRNAs) and microRNAs (miRNAs), have emerged as promising biomarkers of spermatogenic activity (9). CircRNAs are stable, covalently closed RNA molecules that are resistant to exonuclease degradation and exhibit tissue-specific expression (10). Functionally, circRNAs can act as competing endogenous RNAs by binding and sequestering miRNAs, thereby regulating downstream target genes (11). Numerous studies have identified complex circRNA-miRNA-mRNA regulatory networks involved in spermatogenesis, many of which are detectable not only in testicular tissue but also in sperm, seminal plasma, and serum (12). Notably, recent biomarker models incorporating circRNAs have shown encouraging predictive accuracy for micro-TESE outcomes, emphasizing the clinical potential of ncRNA-based signatures (13).
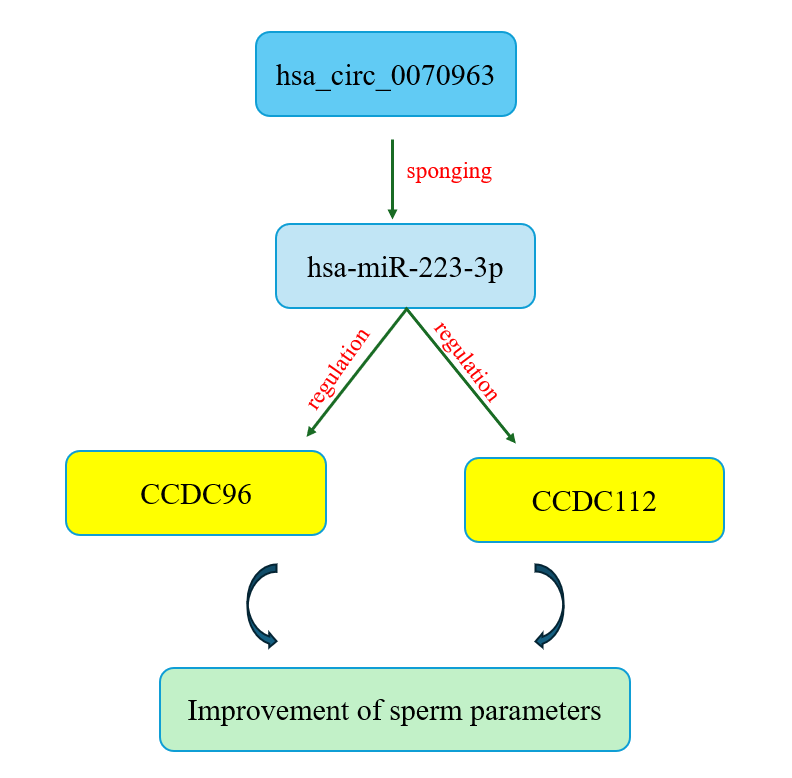
In this context, we identified a novel ceRNA regulatory axis, hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112, through transcriptome reanalysis of testicular biopsies stratified by micro-TESE outcome. Hsa_circ_0070963, an uncharacterized circRNA, was significantly upregulated in NOA individuals with successful sperm retrieval compared to those with failed retrieval. Bioinformatic analyses revealed multiple binding sites for miR-223-3p within hsa_circ_0070963. MiR-223-3p, a miRNA implicated in inflammation and spermatogenic regulation, is predicted to target the mRNAs of CCDC96 and CCDC112, 2 testis-enriched genes markedly downregulated in sperm-negative NOA individuals (14, 15).
CCDC96 and CCDC112 encode proteins essential for sperm flagellar structure and function (16). CCDC112 plays a critical role in mitochondrial sheath formation in the sperm midpiece (17), while CCDC96 is involved in axonemal integrity and ciliary beating (18). Based on these observations, we hypothesized that hsa_circ_0070963 acts as a molecular sponge for miR-223-3p, relieving repression of CCDC96 and CCDC112 and thereby supporting spermatogenesis. To test this hypothesis, we evaluated expression patterns of this axis in blood samples from NOA individuals across different histological subtypes and assessed its diagnostic performance for predicting micro-TESE outcomes relative to existing clinical predictors.
Microsurgical testicular sperm extraction (micro-TESE) has enabled some men with NOA to have their own biological children (6); however, its success rate is limited to roughly 50% (7). Moreover, currently used clinical predictors of sperm retrieval success, such as testicular volume, serum follicle-stimulating hormone (FSH) or inhibin levels, age, and histopathology are inconsistent and unreliable (8). As a result, many individuals undergo an invasive and costly procedure with uncertain outcomes, underscoring the urgent need for reliable, non-invasive biomarkers that can predict micro-TESE success and improve individual counseling and clinical decision-making.
In recent years, non-coding RNAs (ncRNAs), particularly circular RNAs (circRNAs) and microRNAs (miRNAs), have emerged as promising biomarkers of spermatogenic activity (9). CircRNAs are stable, covalently closed RNA molecules that are resistant to exonuclease degradation and exhibit tissue-specific expression (10). Functionally, circRNAs can act as competing endogenous RNAs by binding and sequestering miRNAs, thereby regulating downstream target genes (11). Numerous studies have identified complex circRNA-miRNA-mRNA regulatory networks involved in spermatogenesis, many of which are detectable not only in testicular tissue but also in sperm, seminal plasma, and serum (12). Notably, recent biomarker models incorporating circRNAs have shown encouraging predictive accuracy for micro-TESE outcomes, emphasizing the clinical potential of ncRNA-based signatures (13).
In this context, we identified a novel ceRNA regulatory axis, hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112, through transcriptome reanalysis of testicular biopsies stratified by micro-TESE outcome. Hsa_circ_0070963, an uncharacterized circRNA, was significantly upregulated in NOA individuals with successful sperm retrieval compared to those with failed retrieval. Bioinformatic analyses revealed multiple binding sites for miR-223-3p within hsa_circ_0070963. MiR-223-3p, a miRNA implicated in inflammation and spermatogenic regulation, is predicted to target the mRNAs of CCDC96 and CCDC112, 2 testis-enriched genes markedly downregulated in sperm-negative NOA individuals (14, 15).
CCDC96 and CCDC112 encode proteins essential for sperm flagellar structure and function (16). CCDC112 plays a critical role in mitochondrial sheath formation in the sperm midpiece (17), while CCDC96 is involved in axonemal integrity and ciliary beating (18). Based on these observations, we hypothesized that hsa_circ_0070963 acts as a molecular sponge for miR-223-3p, relieving repression of CCDC96 and CCDC112 and thereby supporting spermatogenesis. To test this hypothesis, we evaluated expression patterns of this axis in blood samples from NOA individuals across different histological subtypes and assessed its diagnostic performance for predicting micro-TESE outcomes relative to existing clinical predictors.
2. Materials and Methods
2.1. Selection of transcriptomic datasets
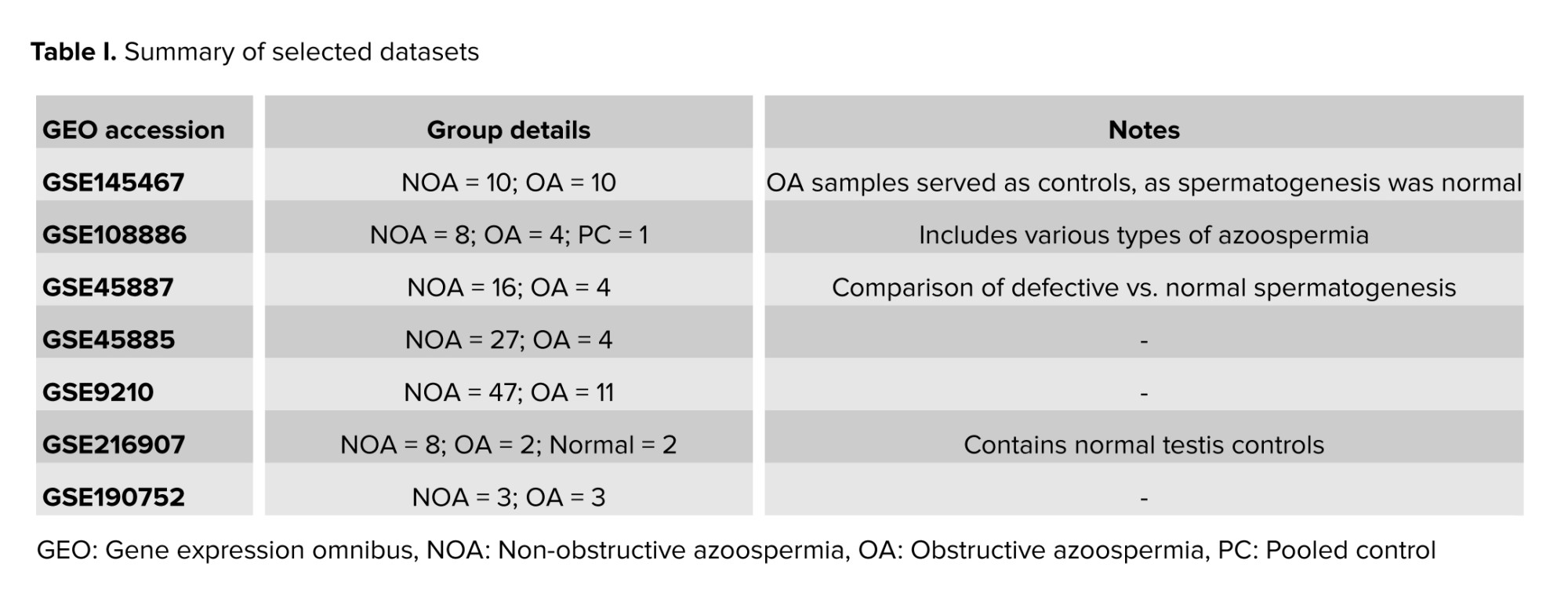
In this case-control study, we performed a systematic search of the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/) to identify transcriptomic datasets reporting differentially expressed genes (DEGs) in testicular tissue from men with impaired spermatogenesis, including NOA. To ensure data quality and reduce potential bias, only studies that met the following criteria were considered: a) human-derived samples only; b) availability of microarray-based gene expression profiles for both infertile individuals and healthy controls; and c) at least 3 independent biological replicates per group. Of the ~35 publicly available datasets on male infertility, 7 met these requirements and were selected for downstream analyses (Table I).
2.2. Identification of DEGs and functional enrichment
Differential expression analysis was performed for each selected data set using the GEO2R platform. Genes that showed an absolute log₂-fold change of more than 2 with a p < 0.01 were considered significantly differentially expressed. To highlight recurrent molecular changes, overlapping DEGs were identified in different data sets. The functional enrichment of these shared genes was then assessed using Enrichr, incorporating both Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways to elucidate the biological processes and signaling networks potentially involved in impaired spermatogenesis.
2.3. Selection of hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112
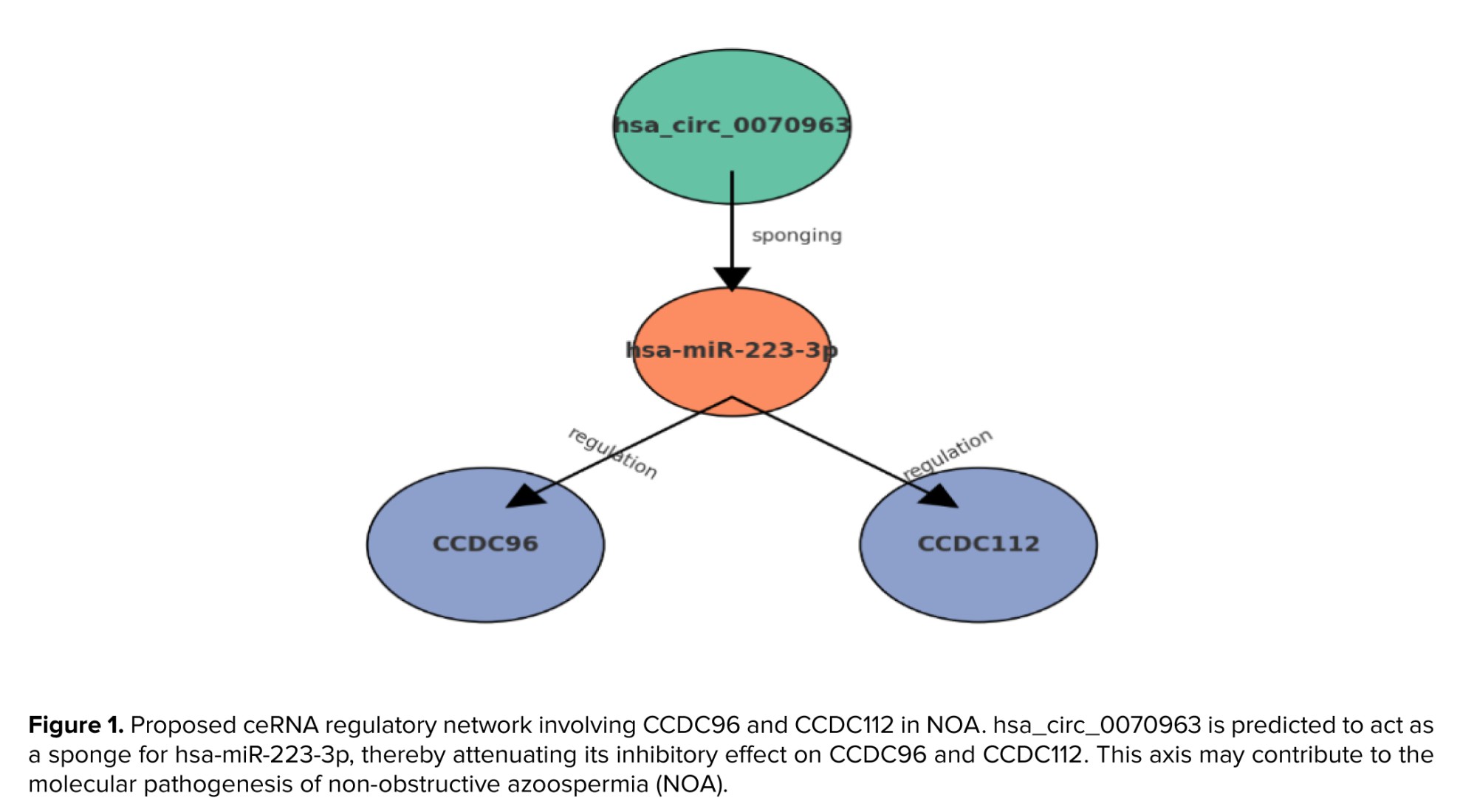
As shown in figure 1, CCDC96 and CCDC112 were found to be the most important downregulated genes in NOA. To investigate the potential post-transcriptional regulation of these genes, we utilized several microRNA prediction resources, including miRTarBase, TargetScan, miRBase, and miRDB. This integrative analysis identified hsa-miR-223-3p as a strong candidate for the regulation of CCDC96 and CCDC112. To further investigate the interactions between competing endogenous RNAs, circRNA databases such as circBank, circBase, CircInteractome, and circAtlas were queried. This search revealed that hsa_circ_0070963 is a putative circRNA sponge capable of sequestering hsa-miR-223-3p and thereby modulating its regulatory activity. Taken together, these results suggest the existence of a novel ceRNA axis: hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112, which may contribute to the molecular pathogenesis of NOA and represents a promising target for further experimental validation.
2.4. Participants
From July to September 2025, a total of 60 men diagnosed with NOA were recruited from the Abortion Research Center of the Reproductive Science Institute in Yazd, Iran. All participants underwent bilateral microTESE as part of assisted reproductive treatment for intracytoplasmic sperm injection.
Before surgery, each individual underwent a comprehensive clinical examination that included standard karyotyping, screening for Y chromosome microdeletions, and measurement of FSH, luteinizing hormone (LH), and serum testosterone.
Individuals with a history of cryptorchidism, previous TESE procedures, hormone therapy, secondary infertility or chromosomal abnormalities, cystic fibrosis, or Y-chromosome microdeletions were excluded. In microTESE, testicular tissue biopsies were taken, fixed in Bouin’s solution, and stained with hematoxylin and eosin for histopathological assessment. Based on established criteria, a specialized pathologist classified the NOA cases as SCOS, HYPO, or MA (n = 20/each). Peripheral blood samples (~3 ml) were also taken from each individual for subsequent molecular analyses. In parallel, 40 fertile men (partners of women with confirmed female infertility) were recruited as a control group. Depending on the results of the microTESE, the NOA individuals were divided into NOA⁺ (successful sperm retrieval; SCOS: n = 3, HYPO: n = 16, MA: n = 10) and NOA⁻ (failed retrieval; n = 31), as summarized in table II.
2.5. RNA extraction, complementary DNA synthesis, and real time-quantitative polymerase chain reaction
After collection of peripheral blood and plasma separation, total RNA was extracted using TRIzol reagent according to the manufacturer’s instructions. RNA concentration and purity were determined using NanoDrop spectrophotometer, and integrity was verified by agarose gel electrophoresis. Equal amounts of high-quality RNA were used for the synthesis of complementary DNA.
To enable simultaneous profiling of miRNAs, circRNAs, and mRNAs while minimizing experimental cost and complexity, we adopted a 2-step strategy. In the first step, miRNAs were polyadenylated in a 20 µL reaction containing 2 µL poly(A) polymerase, 2 µL reaction buffer, 2 µL ATP, 8 µL RNA template, 6 µL nuclease-free water, and incubated at 40°C for 35 min. In the second step, 10 µL of the polyadenylated RNA was subjected to reverse transcription in a 25 µL reaction containing 2 µL reverse transcriptase, 5 µL reverse transcriptase buffer, 1 µL dNTPs, 2.5 µL random hexamers, 1 µL oligo(dT) primer, 1.5 µL miRNA-specific primers, 10 µL RNA template, and 3 µL nuclease-free water. Reverse transcription was performed at 25°C for 15 min, followed by 50°C for 60 min, and ended at 80°C for 6 min.
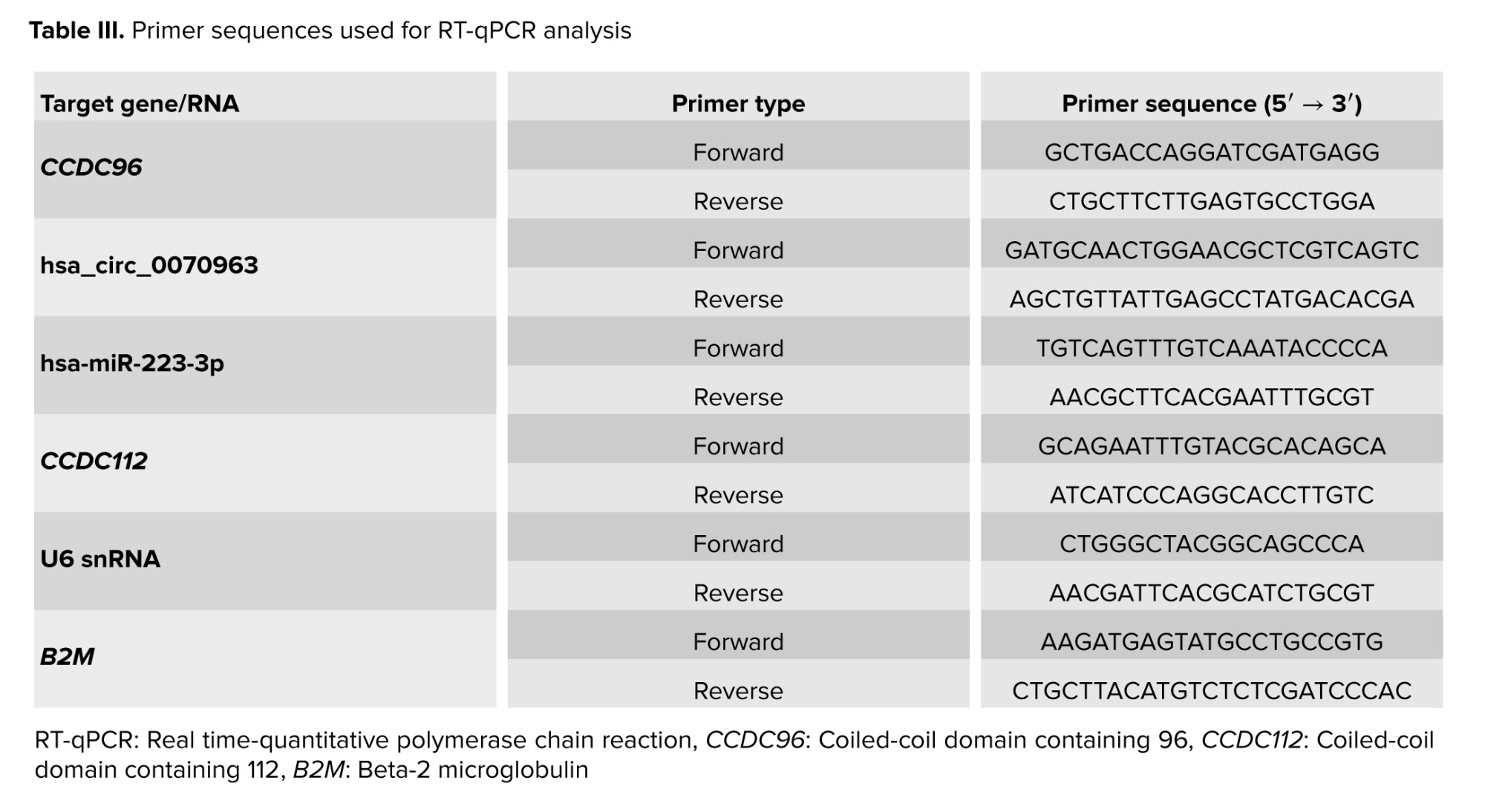
The expression of hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112 was calculated by real-time PCR (Rotor-Gene Q, Qiagen) with specific primers (Table III). Divergent primers spanning the back-splice junction of hsa_circ_0070963 were used to specifically amplify the circular RNA. U6 small nuclear RNA and B2M (β-2-microglobulin) were used as internal reference controls. Relative expression values were calculated using the 2 (-ΔΔCt) method.

2.6. Ethical Considerations
The study protocol was approved by the ethics committee of Shahid Beheshti University of Medical Sciences of Tehran, Iran (Code: IR.SBMU.MSP.REC.1403.331), and written informed consent was obtained from all participants.
2.7. Statistical Analysis
All statistical analyses were performed with GraphPad Prism (version 10). Differences between multiple groups were assessed using one-way analysis of variance (ANOVA) followed by Dunnett’s post hoc test. Comparisons between 2 independent subgroups (NOA⁺ vs. NOA⁻) were performed using an unpaired 2-tailed Student’s t test. A p < 0.05 was considered statistically significant. The diagnostic performance of the biomarker candidates was assessed using receiver operating characteristic (ROC) curve analysis, and the area under the curve (AUC) was calculated to determine their discriminatory ability between NOA⁺ and NOA⁻ individuals.
In this case-control study, we performed a systematic search of the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/) to identify transcriptomic datasets reporting differentially expressed genes (DEGs) in testicular tissue from men with impaired spermatogenesis, including NOA. To ensure data quality and reduce potential bias, only studies that met the following criteria were considered: a) human-derived samples only; b) availability of microarray-based gene expression profiles for both infertile individuals and healthy controls; and c) at least 3 independent biological replicates per group. Of the ~35 publicly available datasets on male infertility, 7 met these requirements and were selected for downstream analyses (Table I).
2.2. Identification of DEGs and functional enrichment
Differential expression analysis was performed for each selected data set using the GEO2R platform. Genes that showed an absolute log₂-fold change of more than 2 with a p < 0.01 were considered significantly differentially expressed. To highlight recurrent molecular changes, overlapping DEGs were identified in different data sets. The functional enrichment of these shared genes was then assessed using Enrichr, incorporating both Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways to elucidate the biological processes and signaling networks potentially involved in impaired spermatogenesis.
2.3. Selection of hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112
As shown in figure 1, CCDC96 and CCDC112 were found to be the most important downregulated genes in NOA. To investigate the potential post-transcriptional regulation of these genes, we utilized several microRNA prediction resources, including miRTarBase, TargetScan, miRBase, and miRDB. This integrative analysis identified hsa-miR-223-3p as a strong candidate for the regulation of CCDC96 and CCDC112. To further investigate the interactions between competing endogenous RNAs, circRNA databases such as circBank, circBase, CircInteractome, and circAtlas were queried. This search revealed that hsa_circ_0070963 is a putative circRNA sponge capable of sequestering hsa-miR-223-3p and thereby modulating its regulatory activity. Taken together, these results suggest the existence of a novel ceRNA axis: hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112, which may contribute to the molecular pathogenesis of NOA and represents a promising target for further experimental validation.
2.4. Participants
From July to September 2025, a total of 60 men diagnosed with NOA were recruited from the Abortion Research Center of the Reproductive Science Institute in Yazd, Iran. All participants underwent bilateral microTESE as part of assisted reproductive treatment for intracytoplasmic sperm injection.
Before surgery, each individual underwent a comprehensive clinical examination that included standard karyotyping, screening for Y chromosome microdeletions, and measurement of FSH, luteinizing hormone (LH), and serum testosterone.
Individuals with a history of cryptorchidism, previous TESE procedures, hormone therapy, secondary infertility or chromosomal abnormalities, cystic fibrosis, or Y-chromosome microdeletions were excluded. In microTESE, testicular tissue biopsies were taken, fixed in Bouin’s solution, and stained with hematoxylin and eosin for histopathological assessment. Based on established criteria, a specialized pathologist classified the NOA cases as SCOS, HYPO, or MA (n = 20/each). Peripheral blood samples (~3 ml) were also taken from each individual for subsequent molecular analyses. In parallel, 40 fertile men (partners of women with confirmed female infertility) were recruited as a control group. Depending on the results of the microTESE, the NOA individuals were divided into NOA⁺ (successful sperm retrieval; SCOS: n = 3, HYPO: n = 16, MA: n = 10) and NOA⁻ (failed retrieval; n = 31), as summarized in table II.
2.5. RNA extraction, complementary DNA synthesis, and real time-quantitative polymerase chain reaction
After collection of peripheral blood and plasma separation, total RNA was extracted using TRIzol reagent according to the manufacturer’s instructions. RNA concentration and purity were determined using NanoDrop spectrophotometer, and integrity was verified by agarose gel electrophoresis. Equal amounts of high-quality RNA were used for the synthesis of complementary DNA.
To enable simultaneous profiling of miRNAs, circRNAs, and mRNAs while minimizing experimental cost and complexity, we adopted a 2-step strategy. In the first step, miRNAs were polyadenylated in a 20 µL reaction containing 2 µL poly(A) polymerase, 2 µL reaction buffer, 2 µL ATP, 8 µL RNA template, 6 µL nuclease-free water, and incubated at 40°C for 35 min. In the second step, 10 µL of the polyadenylated RNA was subjected to reverse transcription in a 25 µL reaction containing 2 µL reverse transcriptase, 5 µL reverse transcriptase buffer, 1 µL dNTPs, 2.5 µL random hexamers, 1 µL oligo(dT) primer, 1.5 µL miRNA-specific primers, 10 µL RNA template, and 3 µL nuclease-free water. Reverse transcription was performed at 25°C for 15 min, followed by 50°C for 60 min, and ended at 80°C for 6 min.
The expression of hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112 was calculated by real-time PCR (Rotor-Gene Q, Qiagen) with specific primers (Table III). Divergent primers spanning the back-splice junction of hsa_circ_0070963 were used to specifically amplify the circular RNA. U6 small nuclear RNA and B2M (β-2-microglobulin) were used as internal reference controls. Relative expression values were calculated using the 2 (-ΔΔCt) method.
2.6. Ethical Considerations
The study protocol was approved by the ethics committee of Shahid Beheshti University of Medical Sciences of Tehran, Iran (Code: IR.SBMU.MSP.REC.1403.331), and written informed consent was obtained from all participants.
2.7. Statistical Analysis
All statistical analyses were performed with GraphPad Prism (version 10). Differences between multiple groups were assessed using one-way analysis of variance (ANOVA) followed by Dunnett’s post hoc test. Comparisons between 2 independent subgroups (NOA⁺ vs. NOA⁻) were performed using an unpaired 2-tailed Student’s t test. A p < 0.05 was considered statistically significant. The diagnostic performance of the biomarker candidates was assessed using receiver operating characteristic (ROC) curve analysis, and the area under the curve (AUC) was calculated to determine their discriminatory ability between NOA⁺ and NOA⁻ individuals.
3. Results
3.1. GO and KEGG pathway enrichment analysis
Analysis of the integrated GEO datasets (Table I) identified a subset of DEGs that were consistently downregulated in NOA. Among these, CCDC96 and CCDC112 were selected for further investigation due to their recurrent dysregulation in several datasets (Figure 2). To complement the functional analysis, we examined their tissue-specific expression patterns using transcriptome data from the Human Protein Atlas and the GTEx project (Figures 1, and 2). Both CCDC96 and CCDC112 showed highly testis-enriched expression with minimal levels detected in non-reproductive tissues, supporting their potential classification as testis-specific genes with critical roles in spermatogenesis.
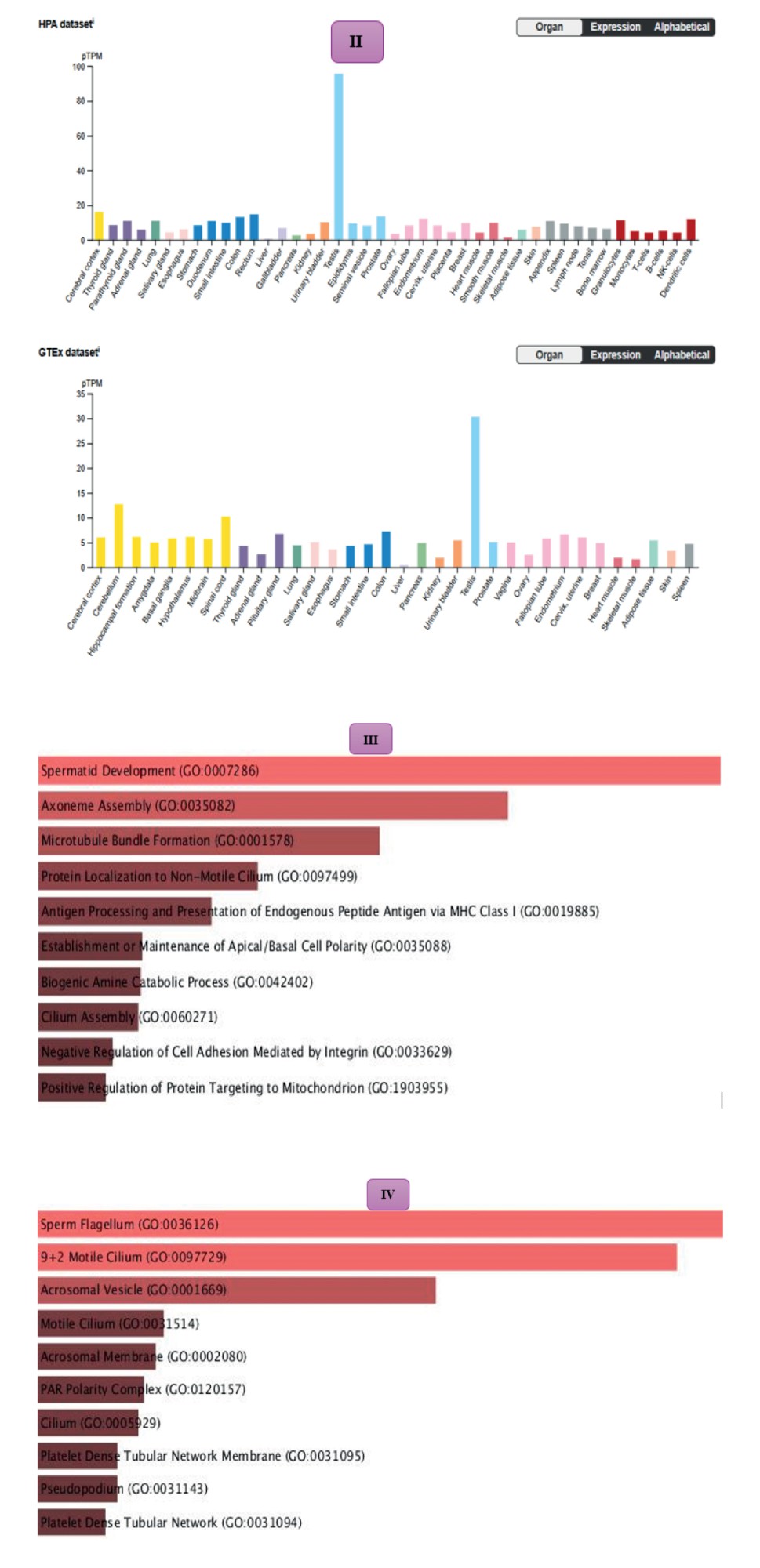
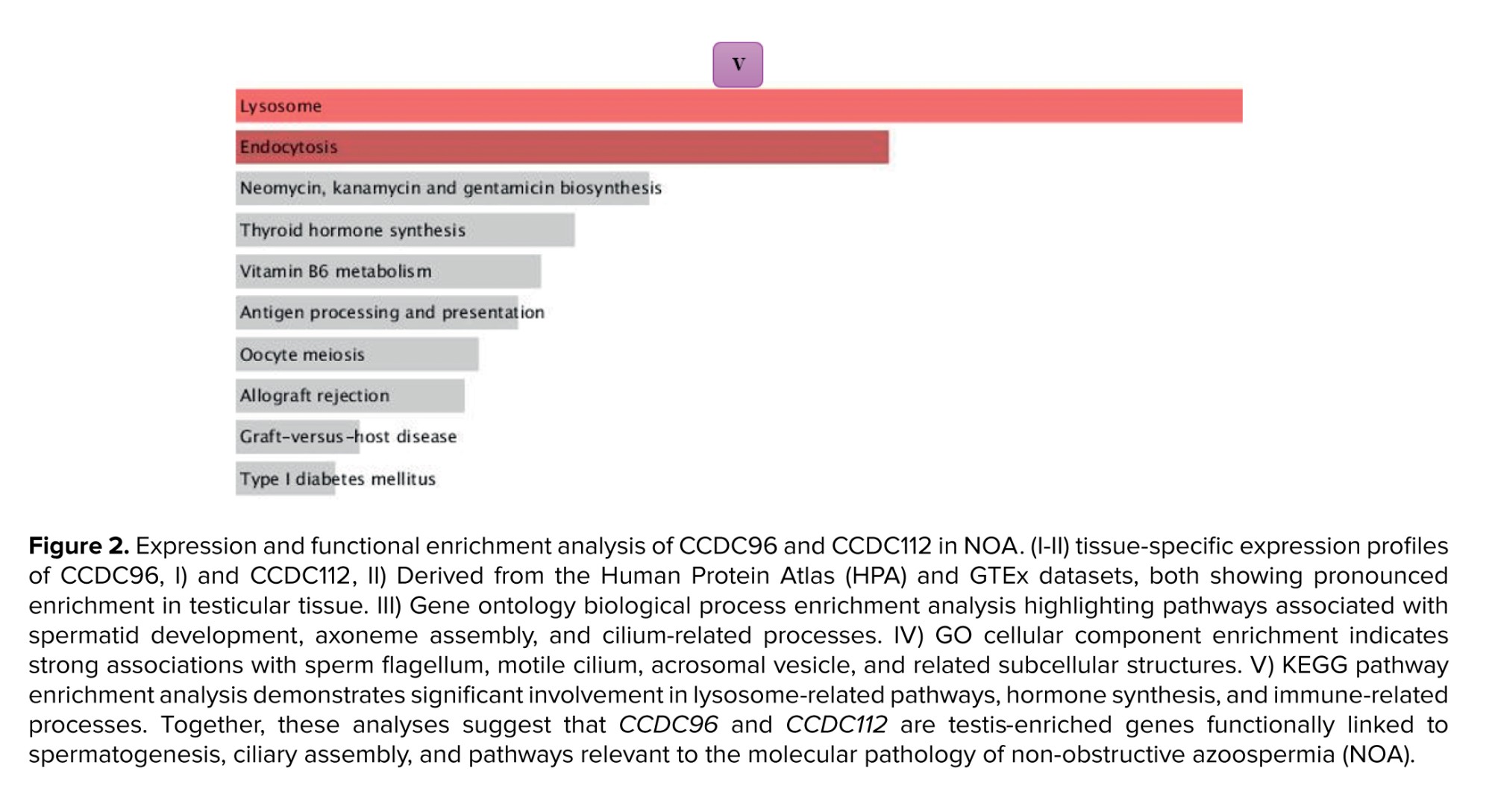
GO and KEGG enrichment analyses were then performed to investigate the biological relevance of the downregulated DEGs (Figures 3-5). Within the biological process category (Figure 3), the most enriched terms were associated with spermatid development, axoneme organization, and cilia assembly, highlighting the importance of these genes for sperm development and flagellar function. In the cellular components domain (Figure 4), the downregulated DEGs were significantly enriched in structural elements central to sperm motility and fertilization, including the sperm flagellum, motile cilium, acrosomal vesicle, and polarity complexes. Finally, analysis of the KEGG pathways (Figure 5) revealed enrichment in lysosome-related processes, hormone metabolism, and immune regulation, reflecting the essential contribution of these pathways to testicular homeostasis and germ cell maturation.
3.2. Clinical features of individuals
Serum concentrations of LH, FSH, and testosterone were measured to assess the baseline hormonal profiles of the study groups. As summarized in table IV, minor differences were observed between the control group, the HYPO, MA, and SCOS; however, none of these differences reached statistical significance (p > 0.05 for all comparisons).
3.3. Gene expression analysis of hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112 in NOA subtypes
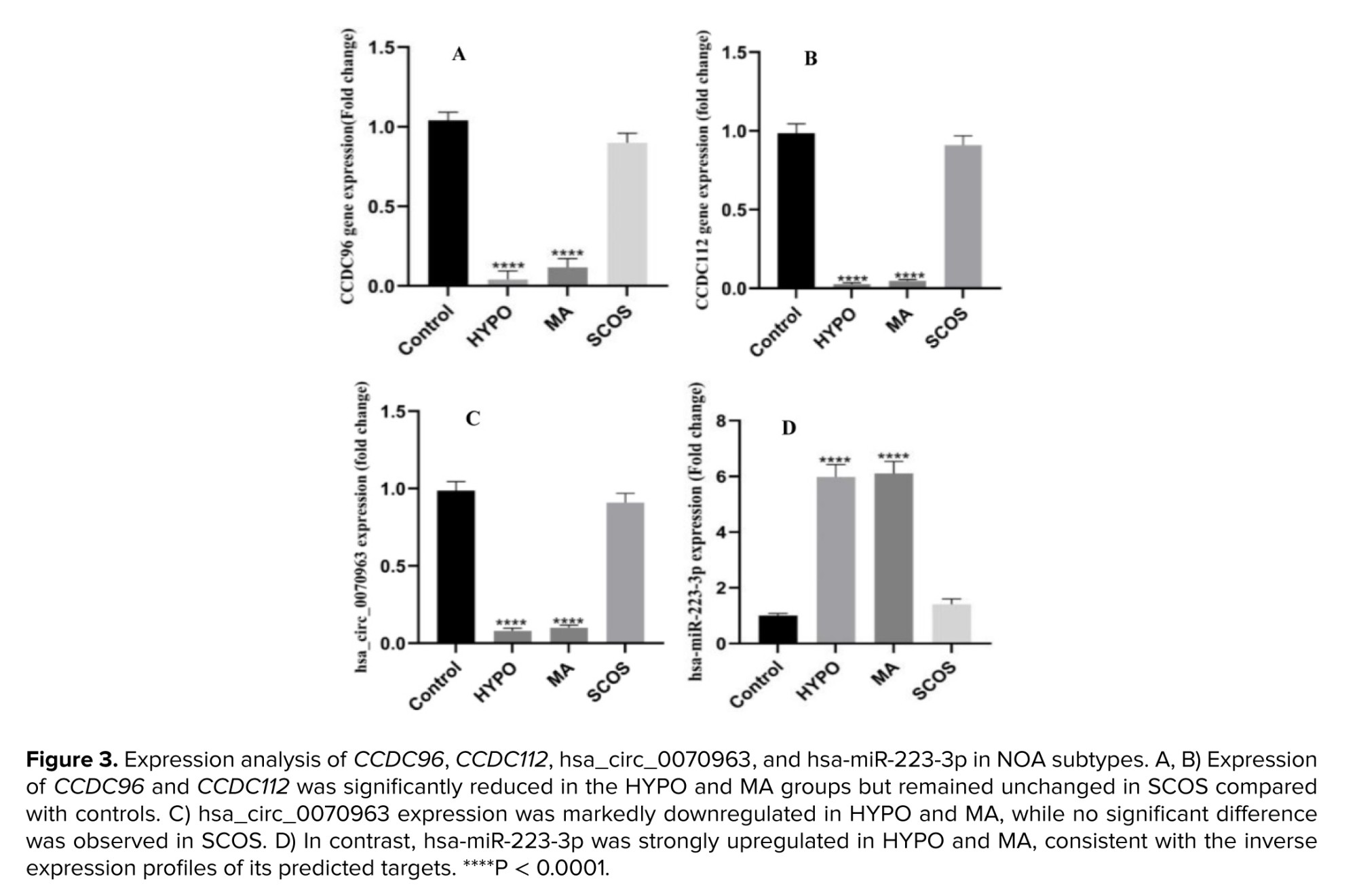
To experimentally validate the bioinformatic predictions, we measured the blood plasma expression levels of CCDC96, CCDC112, their common regulatory miRNA (hsa-miR-223-3p), and the predicted circRNA sponge (hsa_circ_0070963) in individuals with different NOA subtypes (HYPO, MA, SCOS) and healthy controls. As shown in figure 3a, b, the expression of CCDC96 and CCDC112 was significantly downregulated in both HYPO and MA groups (p < 0.0001), while no significant reduction was observed in SCOS individuals. Consistent with these results, hsa_circ_0070963 levels were also significantly reduced in HYPO and MA but remained comparable to controls in SCOS (Figure 3c). In contrast, the upstream regulator hsa-miR-223-3p was strongly upregulated in HYPO and MA subtypes (p < 0.0001), with only modest changes in SCOS (Figure 3d). These reciprocal expression patterns suggest that disruption of the hsa_circ_0070963/hsa-miR-223-3p/CCDC96–CCDC112 regulatory axis may underlie impaired spermatogenesis specifically in HYPO and MA, but not in SCOS, highlighting subtype-specific molecular mechanisms in NOA.
3.4. Predictive value of the hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 axis for mTESE outcomes in NOA individuals
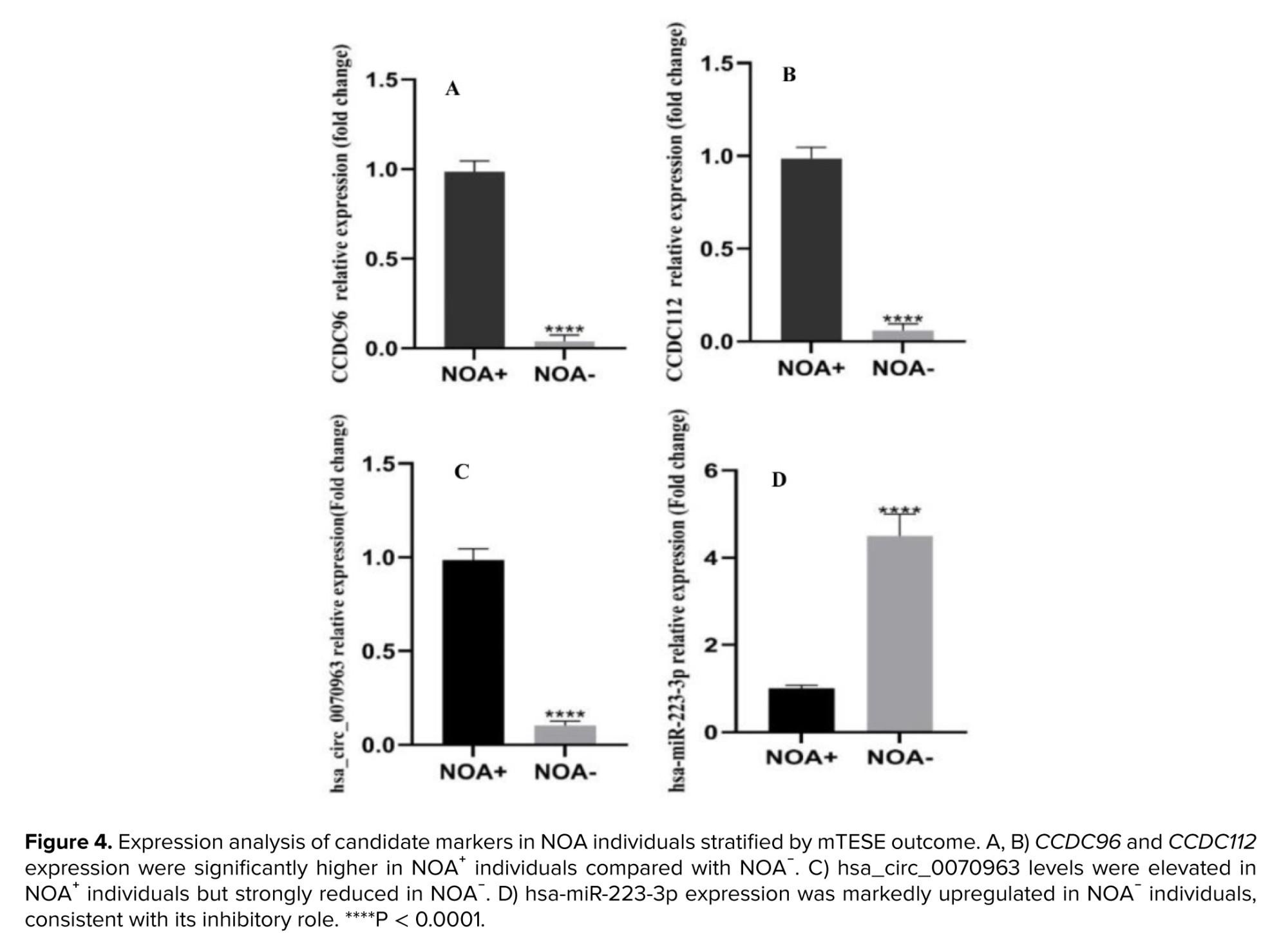
To evaluate whether candidate gene and ncRNA expression could predict sperm retrieval outcomes, we compared NOA individuals with NOA⁺ and NOA⁻ mTESE results. As shown in figure 4a, b, mRNA expression of CCDC96 and CCDC112 were significantly higher in the NOA⁺ group, whereas both genes were profoundly downregulated in NOA⁻ individuals (p < 0.0001). Similarly, expression of the predicted circRNA regulator hsa_circ_0070963 was markedly elevated in NOA⁺ compared to NOA⁻ individuals (Figure 4c). In contrast, the hsa-miR-223-3p displayed the opposite pattern, with significant overexpression in NOA⁻ individuals relative to NOA⁺ (Figure 4d). These reciprocal patterns suggest that the hsa_circ_0070963/hsa-miR-223-3p/CCDC96–CCDC112 regulatory axis plays a critical role in determining spermatogenic capacity and may serve as a predictive biomarker of mTESE outcome in NOA.
3.5. Diagnostic performance of the hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 axis in predicting mTESE outcomes
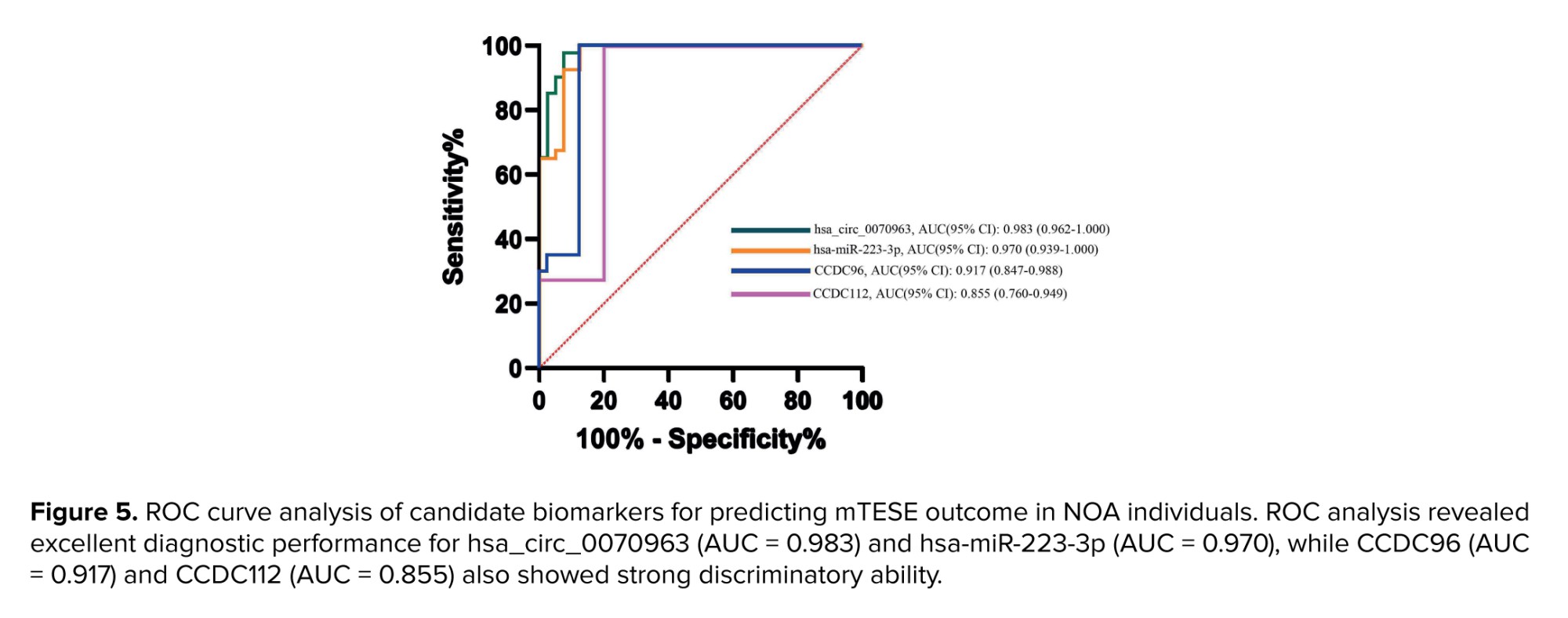
To determine the diagnostic utility of the candidate regulatory components, we performed ROC curve analyses to evaluate their ability to discriminate between NOA⁺ and NOA- individuals (Figure 5). Among the markers tested, hsa_circ_0070963 achieved the highest diagnostic performance with an AUC of 0.983 (95% CI: 0.962-1.000), indicating excellent predictive accuracy. hsa-miR-223-3p also showed strong diagnostic potential (AUC = 0.970, 95% CI: 0.939-1.000), consistent with its inverse association with sperm recovery outcomes. Both CCDC96 (AUC = 0.917, 95% CI: 0.847–0.988) and CCDC112 (AUC = 0.855, 95% CI: 0.760-0.949) showed good discriminatory power, albeit slightly lower than the ncRNAs. Collectively, these results suggest that the hsa_circ_0070963/miR-223-3p/CCDC96-CCDC112 axis holds significant diagnostic value and may serve as a predictive biomarker panel for mTESE outcomes in NOA.
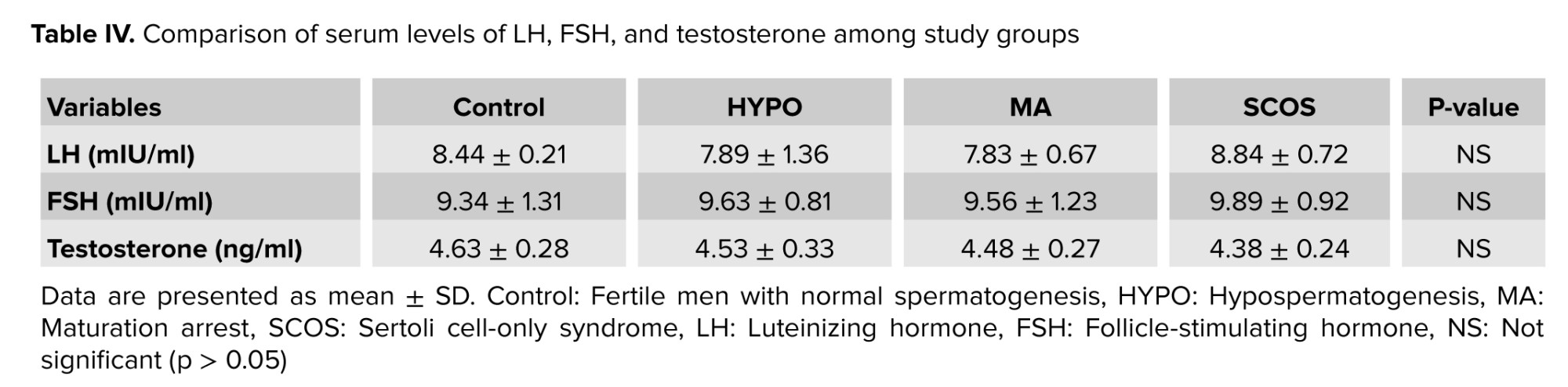
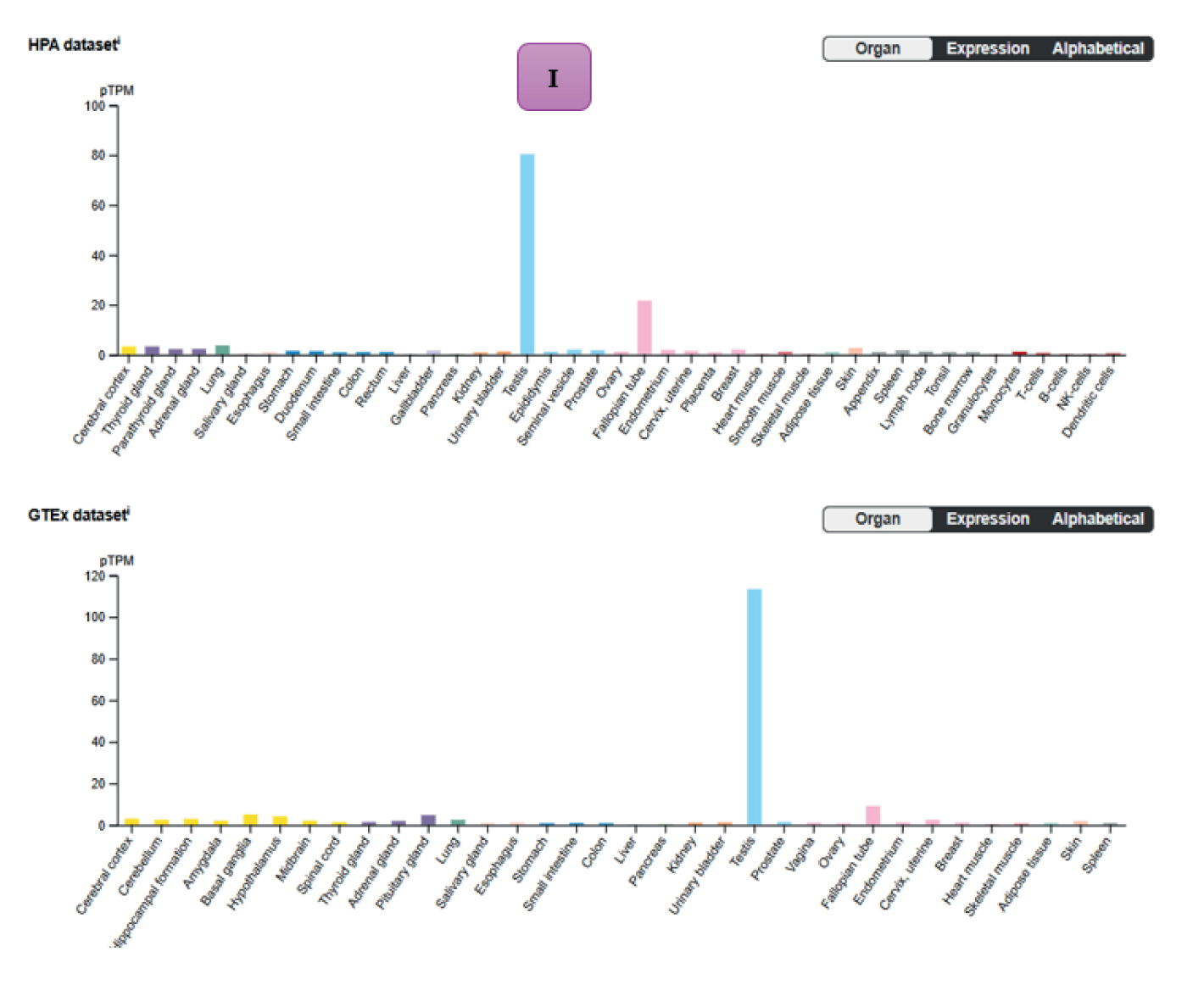
Analysis of the integrated GEO datasets (Table I) identified a subset of DEGs that were consistently downregulated in NOA. Among these, CCDC96 and CCDC112 were selected for further investigation due to their recurrent dysregulation in several datasets (Figure 2). To complement the functional analysis, we examined their tissue-specific expression patterns using transcriptome data from the Human Protein Atlas and the GTEx project (Figures 1, and 2). Both CCDC96 and CCDC112 showed highly testis-enriched expression with minimal levels detected in non-reproductive tissues, supporting their potential classification as testis-specific genes with critical roles in spermatogenesis.
GO and KEGG enrichment analyses were then performed to investigate the biological relevance of the downregulated DEGs (Figures 3-5). Within the biological process category (Figure 3), the most enriched terms were associated with spermatid development, axoneme organization, and cilia assembly, highlighting the importance of these genes for sperm development and flagellar function. In the cellular components domain (Figure 4), the downregulated DEGs were significantly enriched in structural elements central to sperm motility and fertilization, including the sperm flagellum, motile cilium, acrosomal vesicle, and polarity complexes. Finally, analysis of the KEGG pathways (Figure 5) revealed enrichment in lysosome-related processes, hormone metabolism, and immune regulation, reflecting the essential contribution of these pathways to testicular homeostasis and germ cell maturation.
3.2. Clinical features of individuals
Serum concentrations of LH, FSH, and testosterone were measured to assess the baseline hormonal profiles of the study groups. As summarized in table IV, minor differences were observed between the control group, the HYPO, MA, and SCOS; however, none of these differences reached statistical significance (p > 0.05 for all comparisons).
3.3. Gene expression analysis of hsa_circ_0070963/hsa-miR-223-3p/CCDC96 and hsa_circ_0070963/hsa-miR-223-3p/CCDC112 in NOA subtypes
To experimentally validate the bioinformatic predictions, we measured the blood plasma expression levels of CCDC96, CCDC112, their common regulatory miRNA (hsa-miR-223-3p), and the predicted circRNA sponge (hsa_circ_0070963) in individuals with different NOA subtypes (HYPO, MA, SCOS) and healthy controls. As shown in figure 3a, b, the expression of CCDC96 and CCDC112 was significantly downregulated in both HYPO and MA groups (p < 0.0001), while no significant reduction was observed in SCOS individuals. Consistent with these results, hsa_circ_0070963 levels were also significantly reduced in HYPO and MA but remained comparable to controls in SCOS (Figure 3c). In contrast, the upstream regulator hsa-miR-223-3p was strongly upregulated in HYPO and MA subtypes (p < 0.0001), with only modest changes in SCOS (Figure 3d). These reciprocal expression patterns suggest that disruption of the hsa_circ_0070963/hsa-miR-223-3p/CCDC96–CCDC112 regulatory axis may underlie impaired spermatogenesis specifically in HYPO and MA, but not in SCOS, highlighting subtype-specific molecular mechanisms in NOA.
3.4. Predictive value of the hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 axis for mTESE outcomes in NOA individuals
To evaluate whether candidate gene and ncRNA expression could predict sperm retrieval outcomes, we compared NOA individuals with NOA⁺ and NOA⁻ mTESE results. As shown in figure 4a, b, mRNA expression of CCDC96 and CCDC112 were significantly higher in the NOA⁺ group, whereas both genes were profoundly downregulated in NOA⁻ individuals (p < 0.0001). Similarly, expression of the predicted circRNA regulator hsa_circ_0070963 was markedly elevated in NOA⁺ compared to NOA⁻ individuals (Figure 4c). In contrast, the hsa-miR-223-3p displayed the opposite pattern, with significant overexpression in NOA⁻ individuals relative to NOA⁺ (Figure 4d). These reciprocal patterns suggest that the hsa_circ_0070963/hsa-miR-223-3p/CCDC96–CCDC112 regulatory axis plays a critical role in determining spermatogenic capacity and may serve as a predictive biomarker of mTESE outcome in NOA.
3.5. Diagnostic performance of the hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 axis in predicting mTESE outcomes
To determine the diagnostic utility of the candidate regulatory components, we performed ROC curve analyses to evaluate their ability to discriminate between NOA⁺ and NOA- individuals (Figure 5). Among the markers tested, hsa_circ_0070963 achieved the highest diagnostic performance with an AUC of 0.983 (95% CI: 0.962-1.000), indicating excellent predictive accuracy. hsa-miR-223-3p also showed strong diagnostic potential (AUC = 0.970, 95% CI: 0.939-1.000), consistent with its inverse association with sperm recovery outcomes. Both CCDC96 (AUC = 0.917, 95% CI: 0.847–0.988) and CCDC112 (AUC = 0.855, 95% CI: 0.760-0.949) showed good discriminatory power, albeit slightly lower than the ncRNAs. Collectively, these results suggest that the hsa_circ_0070963/miR-223-3p/CCDC96-CCDC112 axis holds significant diagnostic value and may serve as a predictive biomarker panel for mTESE outcomes in NOA.
4. Discussion
Our findings identify the hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 axis as a biologically meaningful and clinically relevant regulatory network in NOA. The coordinated dysregulation of this axis provides new insights into the molecular mechanisms underlying impaired spermatogenesis and highlights its potential utility as both a biomarker and a therapeutic target. Each component of this regulatory pathway plays a distinct yet interconnected role in sperm development, particularly during the late stages of spermiogenesis.
Among the components of this axis, CCDC96 and CCDC112 are of particular importance due to their established structural and functional roles in sperm formation. CCDC112 is a centriolar protein essential for the assembly of the mitochondrial sheath in the sperm midpiece, a structure that supplies the energy required for sperm motility. Experimental ablation of CCDC112 in animal models results in profound defects in mitochondrial organization, compromised flagellar movement, impaired respiration, and ultimately male infertility (19). Similarly, CCDC96 is an integral axonemal protein that forms a functional complex with CCDC113, linking radial spokes to the dynein regulatory complex within the sperm flagellum. Loss of CCDC96 disrupts axonemal integrity and alters ciliary beat frequency and waveform, indicating its role in transmitting mechanical and regulatory signals necessary for proper flagellar motion (16).
These findings align with emerging literature on circRNAs in male infertility. Zhang et al. identified aberrant circRNA profiles in testicular tissue from NOA individuals, suggesting circRNAs could serve as novel biomarkers (9). Liu et al. profiled circRNAs in seminal plasma of infertile men and reported multiple dysregulated circRNAs associated with infertility. They validated a panel of 6 serum circRNAs that predicted sperm retrieval with high accuracy (AUC ≈ 0.977) (12). Our results complement these reports by focusing on one circRNA-miRNA-mRNA axis: we demonstrate that hsa_circ_0070963 and its targets are jointly dysregulated in NOA, and this axis may itself have diagnostic and predictive utility.
These biological roles strongly suggest that adequate expression of CCDC96 and CCDC112 is required for the production of functional, motile spermatozoa. In line with this, our results demonstrate that NOA individuals with successful sperm retrieval (NOA⁺) exhibit significantly higher levels of CCDC96 and CCDC112 mRNA compared with individuals with failed retrieval (NOA⁻). This pattern indicates the presence of residual or focal spermatogenesis in NOA⁺ individuals. In contrast, NOA⁻ individuals- particularly those with SCOS showed near-complete absence of these transcripts, consistent with the lack of germ cells in their testes. Previous transcriptomic studies have also identified CCDC96 as a hub gene distinguishing NOA from normal spermatogenesis, and our data extend these findings by directly linking CCDC96 and CCDC112 expression to sperm retrieval outcomes. Together, these observations support the use of these genes as surrogate molecular markers of spermatogenic potential.
MiR-223-3p represents the inhibitory component of this regulatory axis. Although miR-223 is classically associated with immune cell function, accumulating evidence suggests context-dependent roles in non-immune tissues, including the testis (20). Aberrant expression of miR-223 has been reported in infertile men and in azoospermia-associated miRNA panels, implicating it in testicular dysfunction (15). In our study, miR-223-3p levels were inversely correlated with CCDC96 and CCDC112 expression: NOA⁻ individuals exhibited elevated miR-223-3p levels, whereas NOA⁺ individuals had lower levels. Bioinformatic analyses identified conserved miR-223 binding sites within the 3′ untranslated regions of both CCDC96 and CCDC112, supporting a model in which miR-223-3p directly represses these spermatogenesis-related genes.
The biological significance of miR-223 elevation in severe NOA remains speculative but biologically plausible. One explanation is that miR-223 is enriched in Sertoli cells or infiltrating immune cells in degenerating testes, creating an inflammatory microenvironment that further suppresses germ cell development. Alternatively, persistent expression of miR-223 in germ cells may reflect disrupted differentiation programs (21). Analogous to miR-146a-5p, which must be downregulated for spermatogonial differentiation sustained miR-223 expression may lock germ cells in an immature or dysfunctional state. These observations position miR-223-3p as a negative regulator of key spermatogenic genes within the NOA testis.
At the upstream level of this regulatory network is hsa_circ_0070963, a circular RNA that appears to modulate the axis by sequestering miR-223-3p. Circ_0070963 was among the most upregulated transcripts in NOA⁺ individuals, and computational predictions revealed multiple miR-223-3p response elements within its sequence. Functional rescue experiments further supported this ceRNA relationship, demonstrating that circ_0070963 overexpression attenuates miR-223-mediated repression of CCDC96 and CCDC112 (22). Notably, this is the first study to implicate circ_0070963 in testicular biology. Interestingly, circ_0070963 has previously been shown to sponge miR-223-3p in liver fibrosis, suggesting that this regulatory interaction is conserved across tissues but exerts tissue-specific functional consequences.
In NOA⁺ individuals, upregulation of circ_0070963 may represent a compensatory mechanism that preserves spermatogenic gene expression by limiting miR-223-3p activity. In contrast, reduced circ_0070963 levels in NOA⁻ individuals eliminate this protective effect, allowing miR-223-3p to suppress CCDC96 and CCDC112 translation. This model explains the coordinated expression patterns observed in our cohort: sperm-positive NOA is characterized by high circ_0070963, low miR-223-3p, and high CCDC96/112, whereas sperm-negative NOA shows the opposite profile.
Importantly, this regulatory axis also reflected histopathological heterogeneity within NOA. SCOS samples exhibited minimal circ_0070963 and CCDC96/112 expression alongside elevated miR-223-3p, consistent with complete germ cell absence. MA samples showed intermediate expression levels, reflecting partial spermatogenic progression. HYPO, despite reduced germ cell numbers, displayed expression profiles most similar to normal spermatogenesis, with high circ_0070963 and detectable CCDC96/112. These subtype-specific patterns suggest that the axis mirrors germ cell content and spermatogenic stage.
From a clinical standpoint, the predictive performance of this axis is particularly compelling. Circ_0070963 alone achieved an AUC of approximately 0.9 for predicting micro-TESE success, outperforming traditional predictors such as FSH or testicular volume (7). Combining circ_0070963 with miR-223-3p or downstream mRNAs further improved diagnostic accuracy and specificity. Individuals with unfavorable molecular profiles were highly likely to experience failed sperm retrieval, highlighting the axis’s potential to guide clinical decision-making and avoid unnecessary surgery.
This high specificity is clinically valuable as it could identify those men in whom micro-TESE is likely to fail, allowing alternative options to be considered without the need for surgery. The sensitivity of the axis for detecting residual spermatogenesis was also good. However, some NOA⁺ individuals did not have the same profile, and micro-TESE can sometimes be successful in unfavorable molecular contexts. Nevertheless, the overall diagnostic accuracy of this circRNA-miRNA signature is very encouraging. It compares favorably with other proposed biomarkers; for example, the 4-miRNA panel of a recent study achieved an AUC value of 0.93, and a circMGLL-based nomogram achieved an AUC value of 0.857 (8); our circ_0070963 axis was in a similar performance range. We also noted that the positive predictive value of our model was improved by including histological subtype as a factor. This suggests that an integrative approach combining molecular biomarkers with pathological or hormonal data may be the most reliable predictive tool, as also found in recent machine learning studies.
Nevertheless, several limitations must be acknowledged. The sample size was moderate, validation in larger cohorts is required. Although our data were consistent with the ceRNA mechanism, we did not perform functional assays to demonstrate direct binding. Future experiments, such as luciferase reporter assays or RNA pull-downs will be necessary to confirm that hsa_circ_0070963 directly binds miR-223-3p and that miR-223-3p directly targets CCDC96/112. Moreover, NOA is a heterogeneous condition, and our findings may be most applicable to idiopathic cases. Future studies integrating this axis with additional biomarkers and machine-learning approaches may yield even more robust predictive models.
Overall, our study supports a network-based view of NOA pathogenesis and establishes the hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 axis as a promising molecular signature for understanding and managing severe male infertility.
Among the components of this axis, CCDC96 and CCDC112 are of particular importance due to their established structural and functional roles in sperm formation. CCDC112 is a centriolar protein essential for the assembly of the mitochondrial sheath in the sperm midpiece, a structure that supplies the energy required for sperm motility. Experimental ablation of CCDC112 in animal models results in profound defects in mitochondrial organization, compromised flagellar movement, impaired respiration, and ultimately male infertility (19). Similarly, CCDC96 is an integral axonemal protein that forms a functional complex with CCDC113, linking radial spokes to the dynein regulatory complex within the sperm flagellum. Loss of CCDC96 disrupts axonemal integrity and alters ciliary beat frequency and waveform, indicating its role in transmitting mechanical and regulatory signals necessary for proper flagellar motion (16).
These findings align with emerging literature on circRNAs in male infertility. Zhang et al. identified aberrant circRNA profiles in testicular tissue from NOA individuals, suggesting circRNAs could serve as novel biomarkers (9). Liu et al. profiled circRNAs in seminal plasma of infertile men and reported multiple dysregulated circRNAs associated with infertility. They validated a panel of 6 serum circRNAs that predicted sperm retrieval with high accuracy (AUC ≈ 0.977) (12). Our results complement these reports by focusing on one circRNA-miRNA-mRNA axis: we demonstrate that hsa_circ_0070963 and its targets are jointly dysregulated in NOA, and this axis may itself have diagnostic and predictive utility.
These biological roles strongly suggest that adequate expression of CCDC96 and CCDC112 is required for the production of functional, motile spermatozoa. In line with this, our results demonstrate that NOA individuals with successful sperm retrieval (NOA⁺) exhibit significantly higher levels of CCDC96 and CCDC112 mRNA compared with individuals with failed retrieval (NOA⁻). This pattern indicates the presence of residual or focal spermatogenesis in NOA⁺ individuals. In contrast, NOA⁻ individuals- particularly those with SCOS showed near-complete absence of these transcripts, consistent with the lack of germ cells in their testes. Previous transcriptomic studies have also identified CCDC96 as a hub gene distinguishing NOA from normal spermatogenesis, and our data extend these findings by directly linking CCDC96 and CCDC112 expression to sperm retrieval outcomes. Together, these observations support the use of these genes as surrogate molecular markers of spermatogenic potential.
MiR-223-3p represents the inhibitory component of this regulatory axis. Although miR-223 is classically associated with immune cell function, accumulating evidence suggests context-dependent roles in non-immune tissues, including the testis (20). Aberrant expression of miR-223 has been reported in infertile men and in azoospermia-associated miRNA panels, implicating it in testicular dysfunction (15). In our study, miR-223-3p levels were inversely correlated with CCDC96 and CCDC112 expression: NOA⁻ individuals exhibited elevated miR-223-3p levels, whereas NOA⁺ individuals had lower levels. Bioinformatic analyses identified conserved miR-223 binding sites within the 3′ untranslated regions of both CCDC96 and CCDC112, supporting a model in which miR-223-3p directly represses these spermatogenesis-related genes.
The biological significance of miR-223 elevation in severe NOA remains speculative but biologically plausible. One explanation is that miR-223 is enriched in Sertoli cells or infiltrating immune cells in degenerating testes, creating an inflammatory microenvironment that further suppresses germ cell development. Alternatively, persistent expression of miR-223 in germ cells may reflect disrupted differentiation programs (21). Analogous to miR-146a-5p, which must be downregulated for spermatogonial differentiation sustained miR-223 expression may lock germ cells in an immature or dysfunctional state. These observations position miR-223-3p as a negative regulator of key spermatogenic genes within the NOA testis.
At the upstream level of this regulatory network is hsa_circ_0070963, a circular RNA that appears to modulate the axis by sequestering miR-223-3p. Circ_0070963 was among the most upregulated transcripts in NOA⁺ individuals, and computational predictions revealed multiple miR-223-3p response elements within its sequence. Functional rescue experiments further supported this ceRNA relationship, demonstrating that circ_0070963 overexpression attenuates miR-223-mediated repression of CCDC96 and CCDC112 (22). Notably, this is the first study to implicate circ_0070963 in testicular biology. Interestingly, circ_0070963 has previously been shown to sponge miR-223-3p in liver fibrosis, suggesting that this regulatory interaction is conserved across tissues but exerts tissue-specific functional consequences.
In NOA⁺ individuals, upregulation of circ_0070963 may represent a compensatory mechanism that preserves spermatogenic gene expression by limiting miR-223-3p activity. In contrast, reduced circ_0070963 levels in NOA⁻ individuals eliminate this protective effect, allowing miR-223-3p to suppress CCDC96 and CCDC112 translation. This model explains the coordinated expression patterns observed in our cohort: sperm-positive NOA is characterized by high circ_0070963, low miR-223-3p, and high CCDC96/112, whereas sperm-negative NOA shows the opposite profile.
Importantly, this regulatory axis also reflected histopathological heterogeneity within NOA. SCOS samples exhibited minimal circ_0070963 and CCDC96/112 expression alongside elevated miR-223-3p, consistent with complete germ cell absence. MA samples showed intermediate expression levels, reflecting partial spermatogenic progression. HYPO, despite reduced germ cell numbers, displayed expression profiles most similar to normal spermatogenesis, with high circ_0070963 and detectable CCDC96/112. These subtype-specific patterns suggest that the axis mirrors germ cell content and spermatogenic stage.
From a clinical standpoint, the predictive performance of this axis is particularly compelling. Circ_0070963 alone achieved an AUC of approximately 0.9 for predicting micro-TESE success, outperforming traditional predictors such as FSH or testicular volume (7). Combining circ_0070963 with miR-223-3p or downstream mRNAs further improved diagnostic accuracy and specificity. Individuals with unfavorable molecular profiles were highly likely to experience failed sperm retrieval, highlighting the axis’s potential to guide clinical decision-making and avoid unnecessary surgery.
This high specificity is clinically valuable as it could identify those men in whom micro-TESE is likely to fail, allowing alternative options to be considered without the need for surgery. The sensitivity of the axis for detecting residual spermatogenesis was also good. However, some NOA⁺ individuals did not have the same profile, and micro-TESE can sometimes be successful in unfavorable molecular contexts. Nevertheless, the overall diagnostic accuracy of this circRNA-miRNA signature is very encouraging. It compares favorably with other proposed biomarkers; for example, the 4-miRNA panel of a recent study achieved an AUC value of 0.93, and a circMGLL-based nomogram achieved an AUC value of 0.857 (8); our circ_0070963 axis was in a similar performance range. We also noted that the positive predictive value of our model was improved by including histological subtype as a factor. This suggests that an integrative approach combining molecular biomarkers with pathological or hormonal data may be the most reliable predictive tool, as also found in recent machine learning studies.
Nevertheless, several limitations must be acknowledged. The sample size was moderate, validation in larger cohorts is required. Although our data were consistent with the ceRNA mechanism, we did not perform functional assays to demonstrate direct binding. Future experiments, such as luciferase reporter assays or RNA pull-downs will be necessary to confirm that hsa_circ_0070963 directly binds miR-223-3p and that miR-223-3p directly targets CCDC96/112. Moreover, NOA is a heterogeneous condition, and our findings may be most applicable to idiopathic cases. Future studies integrating this axis with additional biomarkers and machine-learning approaches may yield even more robust predictive models.
Overall, our study supports a network-based view of NOA pathogenesis and establishes the hsa_circ_0070963/miR-223-3p/CCDC96–CCDC112 axis as a promising molecular signature for understanding and managing severe male infertility.
5. Conclusion
In conclusion, this study identifies a novel ceRNA axis, hsa_circ_0070963/miR-223/CCDC96–CCDC112, that enhances our understanding of post-transcriptional regulation in spermatogenesis and demonstrates strong potential as a non-invasive biomarker for predicting mTESE outcomes in NOA. This axis distinguishes individuals with residual spermatogenesis from those without, underscoring the clinical promise of circRNA-miRNA-mRNA networks for personalized infertility management. While further validation and mechanistic studies are required, these findings provide a foundation for integrating molecular profiling into NOA diagnosis and treatment, moving toward a precision medicine approach in male infertility.
Data Availability
All raw data are available from the corresponding author on reasonable request.
Author Contributions
E. Babakhanzadeh and S. Ghafouri-Fard designed the study and conducted the research. A. Khodadadian, M. Nazari, M. Mozhdeh and J. Fazeli monitored, evaluated, and analyzed the results of the study. Further, S. Vahidi Mehrgardi, M.H. Jafari, M. Dehghani, and N. Ghasemi reviewed the article. All authors approved the final manuscript and take responsibility for the integrity of the data.
The study was conducted as a collaborative project between 2 institution. Sampling was performed in Abortion Research Center, Yazd, Iran under prof. Ghasemi supervision and advanced test was performed in Shahid Beheshti University, Tehran, Iran under Dr. Ghafouri-Fard supervision.
Acknowledgments
This article has been extracted from Ph.D. thesis Emad Babakhanzadeh in Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran. This study was supported by the Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran (grant no: 43011482). ChatGPT (OpenAI) has been used to correct spelling and grammatical errors in the article.
Conflict of Interest
The authors declare that there is no conflict of interest.
Data Availability
All raw data are available from the corresponding author on reasonable request.
Author Contributions
E. Babakhanzadeh and S. Ghafouri-Fard designed the study and conducted the research. A. Khodadadian, M. Nazari, M. Mozhdeh and J. Fazeli monitored, evaluated, and analyzed the results of the study. Further, S. Vahidi Mehrgardi, M.H. Jafari, M. Dehghani, and N. Ghasemi reviewed the article. All authors approved the final manuscript and take responsibility for the integrity of the data.
The study was conducted as a collaborative project between 2 institution. Sampling was performed in Abortion Research Center, Yazd, Iran under prof. Ghasemi supervision and advanced test was performed in Shahid Beheshti University, Tehran, Iran under Dr. Ghafouri-Fard supervision.
Acknowledgments
This article has been extracted from Ph.D. thesis Emad Babakhanzadeh in Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran. This study was supported by the Department of Medical Genetics, Shahid Beheshti University of Medical Sciences, Tehran, Iran (grant no: 43011482). ChatGPT (OpenAI) has been used to correct spelling and grammatical errors in the article.
Conflict of Interest
The authors declare that there is no conflict of interest.
Type of Study: Original Article |
Subject:
Reproductive Genetics
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
